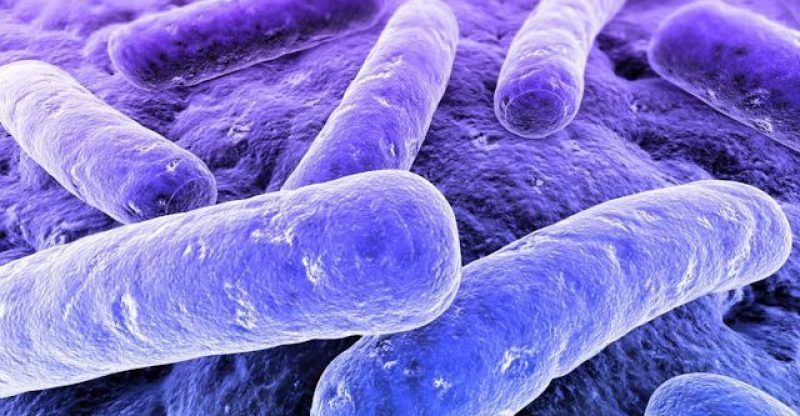
La quimiosíntesis representa uno de los procesos biológicos fundamentales en nuestro planeta, aunque permanece considerablemente menos conocido que su prima famosa, la fotosíntesis. Este fascinante fenómeno metabólico ha revolucionado nuestra comprensión sobre los límites de la vida y ha expandido dramáticamente nuestra percepción acerca de los ambientes donde los organismos pueden prosperar. En este artículo, exploraremos en profundidad el mundo de la quimiosíntesis, desde sus fundamentos bioquímicos hasta su trascendental importancia ecológica y evolutiva.
Definición rápida: La quimiosíntesis es el proceso mediante el cual ciertos microorganismos (principalmente bacterias y arqueas) producen materia orgánica utilizando la energía química obtenida de la oxidación de compuestos inorgánicos, en lugar de usar luz solar como en la fotosíntesis.
Definición y conceptos básicos
La quimiosíntesis es un proceso metabólico mediante el cual ciertos organismos, principalmente bacterias y arqueas, obtienen energía a partir de la oxidación de compuestos inorgánicos en lugar de utilizar la luz solar. Esta energía es posteriormente empleada para sintetizar moléculas orgánicas complejas (como carbohidratos) a partir de dióxido de carbono (CO₂), constituyendo así un mecanismo de fijación de carbono.
La palabra «quimiosíntesis» proviene etimológicamente de «quimio» (relativo a reacciones químicas) y «síntesis» (proceso de formación de compuestos a partir de sustancias más simples). El término fue acuñado por el microbiólogo ruso Sergei Winogradsky a finales del siglo XIX, cuando descubrió bacterias capaces de utilizar compuestos inorgánicos como fuente de energía.
Los organismos quimiosintéticos se clasifican como autótrofos, lo que significa que pueden generar su propio alimento sin depender de otros seres vivos. Sin embargo, a diferencia de los fotoautótrofos (como las plantas), que utilizan la energía lumínica, los quimioautótrofos obtienen su energía de reacciones químicas inorgánicas.
La ecuación general simplificada que describe la quimiosíntesis podría expresarse como:
CO₂ + H₂O + compuesto inorgánico reducido + O₂ → materia orgánica + compuesto inorgánico oxidado
Aunque esta representación es extremadamente simplificada, captura la esencia del proceso: la oxidación de un compuesto inorgánico proporciona la energía necesaria para fijar el dióxido de carbono y sintetizar moléculas orgánicas.
Es importante destacar que, aunque la quimiosíntesis y la fotosíntesis son procesos diferentes en su fuente energética, ambos comparten el objetivo final de fijar carbono inorgánico (CO₂) en moléculas orgánicas. Por esta razón, ambos grupos de organismos son considerados productores primarios en sus respectivos ecosistemas, formando la base de las cadenas tróficas. Sin embargo, mientras que la fotosíntesis sustenta la inmensa mayoría de la vida visible en la superficie del planeta, la quimiosíntesis domina en ambientes extremos donde la luz no penetra, demostrando que la vida ha encontrado múltiples soluciones al desafío fundamental de generar energía metabólica.
Diferencias entre fotosíntesis y quimiosíntesis
| Característica | Fotosíntesis | Quimiosíntesis |
|---|---|---|
| Fuente de energía | Luz solar | Oxidación de compuestos inorgánicos |
| Organismos | Plantas, algas, cianobacterias | Bacterias y arqueas especializadas |
| Ubicación | Ecosistemas iluminados | Ambientes oscuros (fumarolas, suelo, cuevas) |
| Compuestos utilizados | H₂O + CO₂ | H₂S, NH₃, Fe²⁺, CH₄ + CO₂ |
| Productos | Glucosa + O₂ | Glucosa + compuestos oxidados (SO₄²⁻, NO₃⁻) |
| Dependencia de luz | Total | Ninguna |
| Descubrimiento | Conocida desde antigüedad | Winogradsky (1887) |
Historia del descubrimiento
El descubrimiento de la quimiosíntesis constituye uno de los hitos fundamentales en la microbiología y ha transformado radicalmente nuestra comprensión sobre los límites de la vida en la Tierra.
Los primeros indicios: Sergei Winogradsky
El científico ruso Sergei Winogradsky (1856-1953) es considerado el padre de la microbiología del suelo y el descubridor de la quimiosíntesis. Sus investigaciones pioneras en la década de 1880 lo llevaron a observar bacterias con capacidades metabólicas previamente desconocidas.
Durante sus estudios con bacterias del azufre (Beggiatoa), Winogradsky notó que estos microorganismos podían crecer en medios sin materia orgánica, utilizando exclusivamente compuestos inorgánicos como el sulfuro de hidrógeno (H₂S) como fuente de energía. Observó que estas bacterias oxidaban el H₂S a azufre elemental (S) y posteriormente a sulfato (SO₄²⁻), utilizando la energía liberada en estas reacciones para su metabolismo.
En 1887, Winogradsky acuñó el término «quimiosíntesis» para describir este novedoso proceso metabólico, distinguiéndolo claramente de la fotosíntesis. Sus descubrimientos fueron revolucionarios pues, hasta ese momento, se pensaba que todos los autótrofos dependían exclusivamente de la luz solar.

Desarrollos posteriores: siglo XX
Durante la primera mitad del siglo XX, los estudios sobre organismos quimiosintéticos se centraron principalmente en bacterias del suelo y ambientes acuáticos superficiales. Sin embargo, el conocimiento sobre estos procesos dio un salto cualitativo en la década de 1970 con los descubrimientos realizados en las profundidades oceánicas.
En 1977, el descubrimiento de las fumarolas hidrotermales (también conocidas como respiraderos hidrotermales o «vents») en las dorsales oceánicas por el submarino de investigación Alvin reveló ecosistemas sorprendentemente ricos en biodiversidad a pesar de la absoluta oscuridad, las altas presiones y temperaturas extremas. Estos hallazgos desafiaron la concepción tradicional de que todos los ecosistemas dependían ultimamemnte de la energía solar.
Los científicos Holger Jannasch y Colleen Cavanaugh fueron pioneros en demostrar que la base de estas cadenas tróficas eran las bacterias quimiosintéticas, que obtenían energía de la oxidación del sulfuro de hidrógeno emitido por las fumarolas. Este descubrimiento expandió dramáticamente nuestra comprensión sobre la distribución de la vida en el planeta y los límites de la habitabilidad.
En las décadas siguientes, se han identificado numerosos ecosistemas basados en la quimiosíntesis, desde fuentes frías de metano en el fondo oceánico hasta cuevas terrestres ricas en sulfuro, demostrando la extraordinaria importancia ecológica de este proceso metabólico.
Mecanismos bioquímicos
La quimiosíntesis es un proceso metabólico complejo que integra diversas rutas bioquímicas. Aunque existen múltiples variantes según el tipo de organismo y los compuestos inorgánicos utilizados, podemos identificar dos componentes fundamentales en todos los procesos quimiosintéticos:
- Obtención de energía: mediante la oxidación de compuestos inorgánicos reducidos.
- Fijación de carbono: incorporación del CO₂ a moléculas orgánicas utilizando la energía previamente obtenida.
A nivel celular, estos organismos presentan adaptaciones específicas en sus membranas.
Obtención de energía
Los organismos quimiosintéticos obtienen energía a través de reacciones redox (reducción-oxidación) de compuestos inorgánicos. Durante estas reacciones, los electrones son transferidos desde un donador (que se oxida) hasta un aceptor final (que se reduce), generando un gradiente electroquímico a través de la membrana celular.
Este gradiente electroquímico, también conocido como fuerza protón-motriz, es esencialmente idéntico al generado durante la respiración celular aeróbica en mitocondrias, o durante la fase luminosa de la fotosíntesis en cloroplastos. La convergencia evolutiva hacia este mecanismo universal de acoplamiento energético (quimiosmosis) demuestra su eficiencia fundamental. La diferencia radical entre quimiosíntesis y otros procesos radica en la fuente inicial de electrones: mientras que en la respiración provienen de glucosa y en la fotosíntesis del agua, en la quimiosíntesis provienen directamente de compuestos inorgánicos del ambiente geoquímico.
Entre los principales compuestos inorgánicos utilizados como donadores de electrones encontramos:
- Sulfuro de hidrógeno (H₂S): Oxidado a azufre elemental (S⁰) y posteriormente a sulfato (SO₄²⁻). Utilizado por bacterias sulfurosas incoloras y coloradas en fumarolas hidrotermales, sedimentos anóxicos y fuentes termales. Genera aproximadamente 798 kJ/mol.
- Amonio (NH₄⁺): Oxidado primero a nitrito (NO₂⁻) por bacterias como Nitrosomonas (genera
275 kJ/mol), y posteriormente el nitrito es oxidado a nitrato (NO₃⁻) por Nitrobacter (76 kJ/mol). Proceso fundamental en el ciclo del nitrógeno. - Hierro ferroso (Fe²⁺): Oxidado a hierro férrico (Fe³⁺) por bacterias ferruginosas en ambientes ácidos. Genera muy poca energía (~29 kJ/mol a pH 2), requiriendo procesar grandes cantidades de sustrato.
- Hidrógeno molecular (H₂): Oxidado a agua (H₂O) usando diversos aceptores de electrones. Utilizado por bacterias hidrogenotróficas en suelos, sedimentos y fumarolas. Genera ~237 kJ/mol con O₂ como aceptor.
- Metano (CH₄): Oxidado por bacterias metanotróficas aerobias a CO₂, o anaerobias en consorcio con arqueas. Crucial para regular emisiones de este potente gas de efecto invernadero. Genera ~818 kJ/mol en condiciones aerobias.
- Manganeso (Mn²⁺): Oxidado a Mn⁴⁺ por ciertas bacterias en ambientes acuáticos. Menos estudiado que otros tipos pero importante en ciclos biogeoquímicos de lagos y océanos.
- Tiosulfato (S₂O₃²⁻) y otros compuestos de azufre intermedios: Diversos estados de oxidación del azufre pueden servir como donadores, proporcionando flexibilidad metabólica a bacterias del azufre en ambientes fluctuantes.
Como aceptor final de electrones, la mayoría de los quimiosintéticos aerobios utilizan el oxígeno molecular (O₂), que se reduce a agua (H₂O). Sin embargo, en ambientes anaerobios, pueden emplearse otros aceptores como nitrato (NO₃⁻), sulfato (SO₄²⁻) o dióxido de carbono (CO₂).
El flujo de electrones a través de la cadena transportadora genera un gradiente de protones que impulsa la síntesis de ATP (adenosín trifosfato) mediante ATP sintasa, de manera análoga a la fosforilación oxidativa en la respiración celular. Este ATP proporciona la energía necesaria para la fijación de carbono y otras actividades metabólicas.
Fijación de carbono
Una vez obtenida la energía, los organismos quimiosintéticos la utilizan para incorporar CO₂ atmosférico en moléculas orgánicas mediante un proceso denominado fijación de carbono. Aunque existen seis vías diferentes de fijación de carbono conocidas en la naturaleza, la más común entre los quimiosintéticos es el ciclo de Calvin-Benson-Bassham.
El ciclo de Calvin, también presente en las plantas durante la fotosíntesis, utiliza la enzima RuBisCO (ribulosa-1,5-bisfosfato carboxilasa/oxigenasa) para catalizar la reacción entre CO₂ y ribulosa-1,5-bisfosfato, formando dos moléculas de 3-fosfoglicerato que posteriormente se transforman en glucosa y otros compuestos orgánicos.
Algunos organismos quimiosintéticos, especialmente arqueas, utilizan vías alternativas como:
- Ciclo del ácido tricarboxílico reverso (rTCA): una versión invertida del ciclo de Krebs
- Vía del 3-hidroxipropionato: utilizada por algunas bacterias verdes no sulfurosas
- Ciclo del 3-hidroxipropionato/4-hidroxibutirato: presente en arqueas termófilas
- Ciclo del dicarboxilato/4-hidroxibutirato: otra vía utilizada por arqueas
- Vía de Wood-Ljungdahl: empleada por acetogens y metanógenos
La elección de la vía de fijación de carbono está relacionada con las condiciones ambientales y las adaptaciones evolutivas específicas de cada grupo de organismos.
Ejemplo específico: quimiosíntesis basada en el azufre
Para ilustrar estos procesos, consideremos el caso de las bacterias oxidantes del azufre como Thiobacillus:
- Fase de obtención de energía:
- Oxidación del H₂S a azufre elemental: H₂S + ½O₂ → S⁰ + H₂O
- Oxidación del azufre elemental a sulfato: S⁰ + H₂O + 1½O₂ → SO₄²⁻ + 2H⁺
- La energía liberada se utiliza para generar ATP
- Fase de fijación de carbono:
- Utilización del ATP para impulsar el ciclo de Calvin
- Incorporación de CO₂ a moléculas orgánicas: 6CO₂ + 12H₂O → C₆H₁₂O₆ + 6O₂
Este proceso permite a las bacterias sintetizar todos los compuestos orgánicos necesarios para su crecimiento y reproducción, utilizando exclusivamente fuentes inorgánicas.
Tipos de quimiosíntesis según el compuesto oxidado
La diversidad metabólica de los organismos quimiosintéticos es extraordinaria. Dependiendo del compuesto inorgánico que oxidan para obtener energía, podemos clasificar la quimiosíntesis en varios tipos principales, cada uno realizado por grupos específicos de microorganismos adaptados a diferentes nichos ecológicos.
Quimiosíntesis del azufre: Las bacterias sulfurosas, como Beggiatoa y Thiobacillus, oxidan el sulfuro de hidrógeno (H₂S) presente en ambientes volcánicos, fumarolas hidrotermales y sedimentos anóxicos. Este proceso genera azufre elemental que posteriormente puede oxidarse a sulfato (SO₄²⁻). Estas bacterias son fundamentales en las fumarolas del océano profundo, donde forman la base de ecosistemas enteros que incluyen gusanos tubícolas gigantes, almejas y cangrejos ciegos.
Quimiosíntesis del nitrógeno (nitrificación): Este proceso de dos etapas es crucial para el ciclo del nitrógeno en los suelos. Primero, bacterias como Nitrosomonas oxidan el amonio (NH₄⁺) a nitrito (NO₂⁻). Posteriormente, bacterias del género Nitrobacter oxidan el nitrito a nitrato (NO₃⁻), la forma de nitrógeno más fácilmente asimilable por las plantas. Este proceso es fundamental para la agricultura, aunque el uso excesivo de fertilizantes puede saturar esta capacidad natural.
Quimiosíntesis del hierro: Bacterias ferruginosas como Thiobacillus ferrooxidans oxidan el hierro ferroso (Fe²⁺) a hierro férrico (Fe³⁺) en ambientes ácidos como drenajes mineros. Aunque obtienen relativamente poca energía de esta reacción, su adaptación a pH extremadamente bajos (pH 1-2) les permite prosperar en ambientes donde pocos organismos sobreviven.
Quimiosíntesis del hidrógeno: Algunas bacterias y arqueas pueden oxidar hidrógeno molecular (H₂) utilizando oxígeno, nitrato o sulfato como aceptor de electrones. Estas bacterias se encuentran en suelos, sedimentos y fuentes hidrotermales donde se produce H₂ geológicamente.
Metanotrofia: Aunque técnicamente es una forma de quimiosíntesis, merece mención especial. Las bacterias metanotróficas oxidan metano (CH₄) usando oxígeno, jugando un papel crucial en la regulación de este potente gas de efecto invernadero. Viven en interfaces entre ambientes anóxicos productores de metano y ambientes óxicos, como en humedales, arrozales y filtraciones de metano en el fondo oceánico.
Ejemplos de organismos y ecosistemas quimiosintéticos
Los ecosistemas basados en quimiosíntesis se encuentran entre los más extremos y fascinantes del planeta, demostrando que la vida puede prosperar en condiciones que parecerían completamente inhóspitas.
Ecosistemas de fumarolas hidrotermales: Descubiertos en 1977 en las Galápagos, estos oasis de vida en el fondo oceánico (2.000-4.000 metros de profundidad) están dominados por bacterias quimiosintéticas que forman la base de toda la cadena trófica. Los gusanos tubícolas gigantes (Riftia pachyptila) carecen de sistema digestivo y dependen completamente de bacterias quimiosintéticas simbióticas que habitan en su interior. Estas bacterias oxidan el H₂S de las fumarolas, y el gusano les proporciona protección y acceso a oxígeno. Pueden alcanzar más de 2 metros de longitud y formar agregaciones densas alrededor de las emisiones de agua caliente rica en minerales.
Filtraciones frías de metano: En diversas zonas del fondo oceánico, especialmente en márgenes continentales, el metano emerge lentamente desde el subsuelo. Comunidades de arqueas metanotróficas anaerobias, frecuentemente en consorcio con bacterias sulfatorreductoras, oxidan este metano y sostienen ecosistemas que incluyen mejillones, almejas y gusanos tubícolas especializados. Estos ecosistemas pueden persistir durante siglos en el mismo lugar.
Bacterias nitrificantes del suelo: Aunque menos espectaculares visualmente, las bacterias nitrificantes como Nitrosomonas y Nitrobacter son cruciales para la fertilidad del suelo. Transforman el amonio (procedente de la descomposición de materia orgánica o de fertilizantes) en nitratos que las plantas pueden absorber. Un solo gramo de suelo fértil puede contener entre 10⁴ y 10⁶ bacterias nitrificantes, constituyendo una biomasa colectiva considerable.
Cuevas de Movile (Rumania): Esta cueva, aislada del exterior durante 5,5 millones de años, contiene un ecosistema único basado completamente en quimiosíntesis. Bacterias que oxidan sulfuro de hidrógeno forman alfombras flotantes en las aguas de la cueva, sosteniendo una red trófica que incluye 48 especies de invertebrados terrestres y acuáticos, muchas endémicas y ciegas. La atmósfera de la cueva contiene 100 veces más CO₂ y mucho menos oxígeno que el aire normal.
Drenajes ácidos de minas: Aunque son ambientes contaminados creados por actividad humana, las bacterias del hierro como Acidithiobacillus ferrooxidans prosperan en estas aguas extremadamente ácidas (pH 0,5-3), oxidando hierro y azufre. Aunque representan un problema ambiental serio, también ofrecen oportunidades biotecnológicas para la biorremediación y la biominería.
Importancia ecológica y evolutiva de la quimiosíntesis
La quimiosíntesis no es simplemente una curiosidad biológica marginal; desempeña roles fundamentales en el funcionamiento de la biosfera y ha tenido implicaciones profundas en la historia evolutiva de la vida en la Tierra.
Ciclado de nutrientes globales: Los organismos quimiosintéticos son actores esenciales en los ciclos biogeoquímicos del nitrógeno, azufre, hierro y carbono. Las bacterias nitrificantes procesan anualmente miles de millones de toneladas de nitrógeno en suelos y océanos, convirtiendo formas reducidas en nitratos biodisponibles. Sin este proceso, la productividad de los ecosistemas terrestres y acuáticos colapsaría, ya que el nitrógeno es el nutriente limitante más común para el crecimiento vegetal.
Productividad primaria en ambientes extremos: Aunque la fotosíntesis domina la producción de biomasa global (aproximadamente 100-120 gigatoneladas de carbono anualmente), la quimiosíntesis sostiene ecosistemas completos en ambientes donde la fotosíntesis es imposible. Se estima que la producción primaria quimiosintética en fumarolas hidrotermales y filtraciones frías, aunque localmente intensa, representa solo el 0,02-0,05% de la productividad primaria global. Sin embargo, estos ecosistemas albergan biodiversidad única y endémica de alto valor científico.
Orígenes de la vida: La hipótesis de las fumarolas hidrotermales como cuna de la vida ha ganado considerable apoyo científico en las últimas décadas. Las condiciones en estos ambientes (gradientes químicos, superficies minerales catalíticas, disponibilidad de H₂ y CO₂) podrían haber favorecido la emergencia de las primeras formas de vida quimiosintéticas hace más de 3.800 millones de años, mucho antes de la evolución de la fotosíntesis. Los primeros organismos en la Tierra pudieron haber sido quimioautótrofos que oxidaban hidrógeno o hierro ferroso.
Regulación climática: Las bacterias metanotróficas del suelo, sedimentos acuáticos y océanos juegan un papel crucial pero frecuentemente subestimado en la regulación del metano atmosférico. El metano es un gas de efecto invernadero aproximadamente 25 veces más potente que el CO₂ en un horizonte de 100 años. Se estima que las bacterias metanotróficas oxidan entre el 50-80% del metano producido biológicamente antes de que alcance la atmósfera, actuando como un sumidero natural que modera el cambio climático.
Astrobiología y búsqueda de vida extraterrestre: El descubrimiento de ecosistemas quimiosintéticos ha revolucionado nuestra comprensión sobre la habitabilidad planetaria. La quimiosíntesis no requiere luz solar, solo gradientes químicos, lo que expande dramáticamente los ambientes donde podría existir vida. Lunas heladas como Europa (Júpiter) y Encélado (Saturno), que poseen océanos subsuperficiales y actividad hidrotermal, son candidatos principales para albergar vida quimiosintética. Las misiones espaciales futuras a estos cuerpos buscarán específicamente biomarcadores de metabolismos quimiosintéticos.
Aplicaciones biotecnológicas y futuro de la investigación
El estudio de los organismos quimiosintéticos no solo amplía nuestro conocimiento biológico fundamental, sino que también está generando aplicaciones prácticas en biotecnología, ingeniería ambiental e industria.
Biorremediación de contaminantes: Las bacterias quimiosintéticas ofrecen soluciones prometedoras para la descontaminación de ambientes impactados. Thiobacillus ferrooxidans y especies relacionadas pueden utilizarse para extraer metales pesados de suelos y aguas contaminadas mediante procesos de biolixiviación. Estas bacterias oxidan sulfuros metálicos insolubles (como pirita, calcopirita) convirtiéndolos en sulfatos solubles que pueden recuperarse. La industria minera utiliza cada vez más estas técnicas de biominería para extraer cobre, oro y uranio de minerales de baja ley, reduciendo el impacto ambiental comparado con los métodos tradicionales.
Tratamiento de aguas residuales: Los procesos de nitrificación-desnitrificación basados en bacterias quimiosintéticas son fundamentales en las plantas modernas de tratamiento de aguas residuales. Las bacterias nitrificantes convierten el amonio tóxico en nitratos, que posteriormente son reducidos a nitrógeno gaseoso por bacterias desnitrificantes anaerobias, eliminándolo del agua. Investigaciones recientes en el proceso ANAMMOX (oxidación anaerobia de amonio) han identificado bacterias capaces de realizar este proceso de forma más eficiente energéticamente, reduciendo los costos operativos hasta en un 60%.
Producción de biocombustibles: Algunas bacterias metanotróficas están siendo investigadas para convertir metano (de rellenos sanitarios o producción ganadera) en biocombustibles líquidos o proteínas de célula única para alimentación animal. Dado que el metano es un subproducto abundante de diversos procesos industriales y agrícolas, esta bioconversión podría simultáneamente reducir emisiones de gases de efecto invernadero y generar productos de valor.
Extremófilos y enzimas industriales: Los organismos quimiosintéticos de ambientes extremos (alta temperatura, acidez o presión) producen enzimas (extremozimas) con propiedades únicas. Enzimas termoestables de bacterias de fumarolas hidrotermales se utilizan en procesos industriales que requieren altas temperaturas, desde la reacción en cadena de la polimerasa (PCR) en biología molecular hasta la producción de detergentes. El mercado global de extremozimas superó los 3.000 millones de dólares en 2023.
Biomonitorización ambiental: Las comunidades de bacterias quimiosintéticas, especialmente nitrificantes, son indicadores sensibles de la salud del suelo y la calidad del agua. Cambios en su abundancia o actividad pueden señalar contaminación, eutrofización o degradación del ecosistema antes de que aparezcan síntomas macroscópicos.
Organismos quimiosintéticos
La capacidad de realizar quimiosíntesis está notablemente restringida a procariotas (bacterias y arqueas), no habiéndose documentado entre eucariotas. Sin embargo, muchos animales y protistas han desarrollado simbiosis mutualistas con procariotas quimiosintéticos, ampliando indirectamente la distribución de este metabolismo.
Bacterias quimiosintéticas
Las bacterias constituyen el grupo más diverso y mejor estudiado de organismos quimiosintéticos. Podemos clasificarlas según el compuesto inorgánico que utilizan como fuente de energía:
Bacterias oxidantes del azufre
Estos microorganismos obtienen energía de la oxidación de compuestos reducidos de azufre:
- Beggiatoa: bacterias filamentosas que oxidan H₂S a azufre elemental, visible como inclusiones intracelulares blancas.
- Thiobacillus: género que incluye especies capaces de oxidar diversos compuestos de azufre en ambientes ácidos.
- Acidithiobacillus ferrooxidans: antes clasificada como Thiobacillus, puede oxidar tanto compuestos de azufre como hierro ferroso, siendo fundamental en la biolixiviación minera.
- Thiomicrospira: bacterias marinas comunes en fumarolas hidrotermales.
Bacterias oxidantes del hidrógeno
Utilizan hidrógeno molecular (H₂) como donador de electrones:
- Hydrogenobacter: termófilos que crecen óptimamente a temperaturas entre 70-75°C.
- Hydrogenovibrio: género marino frecuentemente asociado a hábitats hidrotermales.
- Bacterias knallgas: grupo funcional capaz de crecer aeróbicamente utilizando H₂ como fuente de energía.
Bacterias nitrificantes
Estas bacterias intervienen en el ciclo del nitrógeno oxidando compuestos nitrogenados:
- Bacterias oxidantes del amonio (AOB): transforman amonio (NH₄⁺) en nitrito (NO₂⁻)
- Géneros: Nitrosomonas, Nitrosospira, Nitrosococcus
- Bacterias oxidantes del nitrito (NOB): transforman nitrito (NO₂⁻) en nitrato (NO₃⁻)
- Géneros: Nitrobacter, Nitrospira, Nitrococcus
Bacterias oxidantes del hierro
Obtienen energía de la oxidación del hierro ferroso (Fe²⁺) a férrico (Fe³⁺):
- Gallionella: forma estructuras típicas en forma de «tallos» retorcidos de óxido férrico.
- Leptothrix: produce vainas tubulares impregnadas de hierro y manganeso.
- Mariprofundus ferrooxydans: bacteria marina descubierta recientemente en montes submarinos.
Bacterias oxidantes del metano
También llamadas metanótrofas, utilizan metano como fuente de carbono y energía:
- Methylococcus: cocos gramnegativos con un sistema de membranas intracelulares.
- Methylosinus: bacilos o vibrios provistos de prostecas y reproducción por gemación.
Arqueas quimiosintéticas
Las arqueas representan otro dominio fundamental de organismos quimiosintéticos, destacando por su predominio en ambientes extremos:
Arqueas oxidantes del amonio (AOA)
Descubiertas a principios del siglo XXI, incluyen:
- Nitrosopumilus: primer AOA cultivado en laboratorio, abundante en océanos.
- Nitrososphaera: predominantes en suelos agrícolas.
- Nitrosocaldus: habitan en fuentes termales.
Las investigaciones recientes sugieren que las AOA pueden ser más abundantes que las bacterias nitrificantes en muchos ecosistemas marinos y terrestres, revelando una importancia ecológica previamente subestimada.
Arqueas metanógenas que funcionan reversiblemente
Algunas arqueas metanógenas pueden invertir su metabolismo en determinadas condiciones:
- Methanosarcina: en ciertas condiciones puede oxidar metano acoplándolo a la reducción de nitrato o sulfato.
- Methanothermobacter: puede realizar oxidación de metano en ambientes termófilos.
Arqueas hipertermófilas
Adaptadas a temperaturas extremadamente altas:
- Pyrolobus fumarii: puede crecer a temperaturas de hasta 113°C usando H₂ como donador de electrones.
- Methanopyrus: utiliza H₂ para reducir CO₂ a metano a temperaturas cercanas a 100°C.
Simbiosis quimiosintéticas
Numerosos animales han evolucionado relaciones mutualistas con bacterias quimiosintéticas, permitiéndoles habitar nichos ecológicos inaccesibles a otros organismos heterótrofos:
Gusanos tubícolas gigantes
- Riftia pachyptila: el gusano tubícola gigante carece de aparato digestivo y depende completamente de bacterias quimiosintéticas que alberga en un órgano especializado llamado trofosoma. Estos gusanos pueden alcanzar 2 metros de longitud y habitan exclusivamente cerca de fumarolas hidrotermales.
Bivalvos
- Almejas gigantes (Calyptogena magnifica): albergan bacterias simbiontes en sus branquias.
- Mejillones de fumarolas (Bathymodiolus sp.): mantienen simbiosis duales, albergando tanto bacterias oxidantes del azufre como del metano.
Otros invertebrados con simbiontes quimiosintéticos
- Gusanos siboglinidos: familia que incluye varios grupos con simbiontes quimiosintéticos.
- Camarones (Rimicaris exoculata): cultivan bacterias oxidantes del azufre en sus cámaras branquiales.
- Anémonas: algunas especies de aguas profundas dependen parcialmente de simbiosis quimiosintéticas.

Ecología de la quimiosíntesis
La quimiosíntesis sustenta ecosistemas enteros en ambientes donde la fotosíntesis es imposible debido a la ausencia de luz. Estos ecosistemas quimiosintéticos representan oasis de vida en lo que de otro modo serían desiertos biológicos.
Ecosistemas marinos profundos
Fumarolas hidrotermales (vents)
Las fumarolas hidrotermales son grietas en el lecho marino a través de las cuales fluye agua geotérmicamente calentada. Se forman principalmente en las dorsales oceánicas, donde el magma está relativamente cerca de la superficie del fondo marino.
El agua que emana de estas fumarolas puede alcanzar temperaturas de hasta 400°C y está enriquecida con minereales y compuestos químicos como el sulfuro de hidrógeno, metano, hierro y manganeso. Al contacto con el agua fría del océano (aproximadamente 2°C), estos minerales precipitan formando estructuras conocidas como chimeneas hidrotermales o «fumadores negros» (black smokers).
Estos ecosistemas presentan biomasa y biodiversidad sorprendentemente altas, comparables a las de arrecifes de coral, a pesar de desarrollarse en completa oscuridad. Entre sus características ecológicas destacan:
- Rápido crecimiento: algunos organismos como Riftia pachyptila pueden crecer hasta 85 cm por año.
- Alta productividad: las tasas de producción primaria pueden alcanzar valores similares a los de bosques terrestres.
- Endemismo: aproximadamente el 70% de las especies son endémicas de estos ambientes.
- Eficiencia energética: las bacterias quimiosintéticas pueden fijar hasta el 90% del carbono disponible.
Filtraciones frías (cold seeps)
Las filtraciones frías son áreas del lecho marino donde fluidos ricos en hidrocarburos (principalmente metano) se filtran a través del sedimento. A diferencia de las fumarolas hidrotermales, estos fluidos están a la misma temperatura que el agua circundante.
Las comunidades biológicas de las filtraciones frías están dominadas por:
- Tapetes microbianos compuestos por consorcios de arqueas oxidantes anaeróbicas de metano y bacterias reductoras de sulfato.
- Bivalvos de las familias Vesicomyidae y Solemyidae con simbiontes quimiosintéticos.
- Gusanos poliquetos siboglinidos.
Una característica distintiva de estos ecosistemas es la formación de carbonatos autigénicos, rocas formadas por la precipitación de carbonato cálcico como resultado del metabolismo microbiano.
Cadáveres de ballenas (whale falls)
Cuando una ballena muere y su cadáver se deposita en el fondo marino, crea un ecosistema efímero pero extraordinariamente rico. Los científicos han identificado cuatro etapas sucesionales en estos ecosistemas:
- Fase móvil-carroñera: donde tiburones y otros carroñeros consumen tejidos blandos.
- Fase oportunista: dominada por invertebrados que colonizan huesos y sedimentos enriquecidos.
- Fase reductora de sulfato: donde bacterias anaeróbicas descomponen lípidos óseos, produciendo sulfuro de hidrógeno.
- Fase arrecife: caracterizada por la persistencia de comunidades oportunistas en huesos mineralizados.
Durante la fase reductora de sulfato, que puede durar décadas, se desarrolla un ecosistema basado en la quimiosíntesis similar al de fumarolas hidrotermales, con gusanos siboglinidos, bivalvos y crustáceos especializados.
Ecosistemas terrestres y de agua dulce
Cuevas sulfurosas
Algunas cuevas contienen manantiales ricos en sulfuro de hidrógeno que sustentan comunidades microbianas quimiosintéticas:
- Cueva de Villa Luz (México): famosa por sus «snottites» (biofilms microbianos que cuelgan del techo como estalactitas viscosas).
- Cuevas de Movile (Rumania): ecosistema subterráneo aislado durante millones de años con más de 30 especies endémicas.
- Sistema Frasassi (Italia): extenso sistema de cuevas con abundantes comunidades microbianas quimiosintéticas.
Manantiales termales terrestres
En manantiales termales como los del Parque Nacional Yellowstone, prosperan comunidades de bacterias y arqueas quimiosintéticas adaptadas a temperaturas extremas y condiciones geoquímicas específicas.
Acuíferos profundos
Investigaciones recientes han revelado la existencia de una biosfera profunda en acuíferos subterráneos, donde las comunidades microbianas subsisten mediante quimiosíntesis basada principalmente en la oxidación del hidrógeno.

Quimiosíntesis vs. Fotosíntesis
Aunque tanto la quimiosíntesis como la fotosíntesis son procesos autotróficos de fijación de carbono, presentan diferencias fundamentales:
| Característica | Quimiosíntesis | Fotosíntesis |
|---|---|---|
| Fuente de energía | Compuestos químicos inorgánicos reducidos | Luz solar |
| Organismos | Exclusivamente procariotas (bacterias y arqueas) | Plantas, algas, algunas bacterias y arqueas |
| Dependencia de luz | No requiere luz (puede ocurrir en oscuridad total) | Estrictamente dependiente de luz |
| Pigmentos | No utiliza pigmentos fotosintéticos | Clorofilas, carotenoides, ficobilinas |
| Producción de O₂ | Generalmente no produce O₂ (excepto en nitrificación) | Libera O₂ como subproducto |
| Eficiencia energética | Variable según el compuesto oxidado (generalmente menor) | Aproximadamente 1-2% de conversión de energía solar |
| Distribución ecológica | Principalmente ambientes extremos o marginales | Dominante en ecosistemas terrestres y acuáticos superficiales |
| Impacto global | Local, pero crucial en ciertos ecosistemas | Principal responsable de la producción primaria planetaria |
Importancia comparativa
La fotosíntesis constituye la base de aproximadamente el 99% de la productividad primaria global, convirtiendo anualmente unas 100 mil millones de toneladas de carbono en biomasa. En contraste, la quimiosíntesis contribuye con menos del 1% a la productividad total del planeta.
Sin embargo, la importancia de la quimiosíntesis no debe medirse únicamente en términos cuantitativos. Su valor radica en:
- Diversificación metabólica: amplía significativamente la variedad de ambientes donde la vida puede prosperar.
- Ciclos biogeoquímicos: juega papeles críticos en los ciclos del nitrógeno, azufre y carbono.
- Perspectiva evolutiva: probablemente representó la primera forma de autotrofía en la Tierra primitiva.
Importancia evolutiva
La quimiosíntesis no es simplemente una curiosidad biológica confinada a nichos extremos; representa un proceso metabólico con profundas implicaciones para nuestra comprensión del origen y evolución de la vida.
Quimiosíntesis y el origen de la vida
Numerosos científicos proponen que los primeros organismos autótrofos en la Tierra primitiva habrían utilizado la quimiosíntesis, y no la fotosíntesis, como mecanismo metabólico principal. Esta hipótesis se sustenta en diversos argumentos:
- La atmósfera primitiva carecía de oxígeno y probablemente contenía altas concentraciones de gases como H₂, CO₂, CH₄, NH₃ y H₂S, potenciales sustratos para reacciones quimiosintéticas.
- La radiación ultravioleta intensa habría dificultado la vida en la superficie terrestre, favoreciendo el desarrollo de ecosistemas en ambientes acuáticos profundos o subterráneos.
- Las fuentes hidrotermales submarinas, similares a las actuales, habrían proporcionado gradientes químicos y térmicos estables, así como protección frente a condiciones superficiales adversas.
- Los minerales inorgánicos presentes en estas fuentes (como pirita y otros sulfuros metálicos) podrían haber catalizado las primeras reacciones metabólicas.
La hipótesis del mundo de hierro-azufre, propuesta por Günter Wächtershäuser, sugiere que la vida podría haber surgido en superficie minerales de sulfuro de hierro cerca de fumarolas hidrotermales, donde reacciones quimiosintéticas primitivas habrían precedido a la evolución de organismos celulares.
Paralelamente, la teoría del origen hidrotermal defendida por Michael Russell postula que los gradientes electroquímicos naturales en chimeneas alcalinas habrían facilitado las primeras reacciones metabólicas, de manera análoga a como funcionan actualmente las membranas celulares.
Evolución metabólica
La reconstrucción filogenética sugiere que las vías metabólicas quimiosintéticas probablemente precedieron evolutivamente a la fotosíntesis:
- Quimiosíntesis anaerobia primitiva: utilizando H₂ como donador de electrones y CO₂ o sulfato como aceptor.
- Fotosíntesis anoxigénica: aparición de pigmentos capaces de captar energía lumínica sin producir oxígeno.
- Fotosíntesis oxigénica: desarrollo del fotosistema II que permitió utilizar H₂O como donador de electrones, liberando O₂.
- Diversificación de la quimiosíntesis aerobia: aprovechando el O₂ atmosférico como aceptor terminal de electrones.
Este modelo evolutivo explica la paradoja del oxígeno: los organismos aerobios actuales dependen del oxígeno producido por la fotosíntesis, pero la fotosíntesis oxigénica requirió complejas adaptaciones bioquímicas que probablemente evolucionaron a partir de metabolismos quimiosintéticos previos.
Extremófilos y límites de la vida
El estudio de organismos quimiosintéticos ha revolucionado nuestra comprensión sobre los límites físicos y químicos de la vida. Bacterias y arqueas quimiosintéticas han sido encontradas en:
- Aguas con temperaturas cercanas al punto de ebullición (hasta 122°C)
- Ambientes hipersalinos (salmueras con saturación de NaCl)
- Condiciones extremadamente ácidas (pH < 1)
- Profundidades oceánicas con presiones superiores a 1000 atmósferas
- Rocas subterráneas a varios kilómetros de profundidad
Estos descubrimientos han expandido dramáticamente los parámetros considerados compatibles con la vida, con importantes implicaciones para la astrobiología y la búsqueda de vida extraterrestre.
Aplicaciones y relevancia contemporánea
La quimiosíntesis trasciende el ámbito puramente teórico para ofrecer aplicaciones prácticas en diversos campos:
Biotecnología ambiental
Tratamiento de aguas residuales
Las bacterias nitrificantes juegan un papel fundamental en las plantas de tratamiento de aguas residuales, donde transforman el amonio tóxico (proveniente de la descomposición de materia orgánica) en nitrato, menos nocivo y utilizable como nutriente por plantas:
- Nitrificación: NH₄⁺ → NO₂⁻ → NO₃⁻ (realizada por bacterias quimiosintéticas)
- Desnitrificación: NO₃⁻ → N₂ (realizada por bacterias heterótrofas)
Este proceso, conocido como eliminación biológica de nitrógeno, reduce significativamente el potencial eutrofizante de los efluentes.
Biorremediación
Diversos microorganismos quimiosintéticos participan en procesos de biorremediación:
- Bacterias oxidantes del azufre: neutralizan ambientes alcalinos y eliminan sulfuro de hidrógeno tóxico.
- Bacterias ferrooxidantes: inmovilizan metales pesados mediante su oxidación y precipitación.
- Bacterias metanótrofas: pueden degradar diversos contaminantes orgánicos clorados además de oxidar metano, un potente gas de efecto invernadero.
Tratamiento de drenaje ácido de minas
El drenaje ácido de minas (DAM) constituye uno de los problemas ambientales más graves asociados a la industria minera. Caracterizado por pH extremadamente bajo y altas concentraciones de metales disueltos, el DAM puede contaminar cursos de agua durante décadas o incluso siglos.
Las bacterias acidófilas como Acidithiobacillus ferrooxidans son utilizadas en biorreactores sulfidogénicos que promueven:
- Neutralización del pH.
- Precipitación de metales como sulfuros insolubles.
- Reducción de sulfato a sulfuro.
Aplicaciones industriales
Biominería
La biolixiviación utiliza microorganismos quimiosintéticos para extraer metales de menas de baja ley, ofreciendo ventajas significativas frente a métodos tradicionales:
- Menor impacto ambiental (reducción de emisiones de SO₂).
- Viabilidad económica para yacimientos de baja concentración.
- Menor consumo energético.
Aproximadamente el 15% del cobre mundial se obtiene actualmente mediante biolixiviación, principalmente utilizando Acidithiobacillus ferrooxidans y especies relacionadas.
Producción de biocombustibles
Investigaciones recientes exploran el potencial de bacterias metanótrofas para convertir metano en compuestos de mayor valor:
- Metanol: oxidación parcial de metano.
- Bioplásticos: algunas metanótrofas acumulan polihidroxibutirato (PHB) como reserva energética.
- Proteína unicelular: biomasa bacteriana rica en proteínas para alimentación animal.
Esta tecnología podría transformar el metano, frecuentemente quemado como residuo en explotaciones petrolíferas, en productos comercialmente valiosos.
Implicaciones en astrobiología
El descubrimiento de ecosistemas basados en quimiosíntesis ha revolucionado la búsqueda de vida extraterrestre:
Vida en otros cuerpos celestes
Varios cuerpos del Sistema Solar presentan condiciones potencialmente favorables para organismos quimiosintéticos:
- Europa (luna de Júpiter): evidencia de un océano subsuperficial y posibles fuentes hidrotermales.
- Encélado (luna de Saturno): plumas de agua con compuestos orgánicos y evidencia de actividad hidrotermal.
- Marte: posible presencia histórica de aguas subterráneas y evidencia de metano atmosférico.
- Titán (luna de Saturno): lagos de hidrocarburos que podrían sustentar formas de vida basadas en metano.
Instrumentación para misiones espaciales
La comprensión de la quimiosíntesis ha influido en el diseño de instrumentos para misiones astrobiológicas:
- Espectrómetros de masas: para detectar biomarcadores asociados al metabolismo quimiosintético.
- Sensores electroquímicos: capaces de detectar gradientes redox característicos de actividad biológica.
- Microlaboratorios: diseñados para cultivar potenciales microorganismos extraterrestres in situ.

Perspectivas futuras
El campo de estudio de la quimiosíntesis continúa expandiéndose, planteando desafíos y oportunidades:
Retos en investigación básica
- Diversidad metabólica inexplorada: se estima que menos del 1% de los microorganismos ambientales han sido cultivados en laboratorio.
- Interacciones simbióticas complejas: muchas asociaciones entre quimiosintéticos y eucariotas permanecen pobremente caracterizadas.
- Biología de sistemas: integración de datos -ómicos para comprender holísticamente las redes metabólicas quimiosintéticas.
- Biología sintética: diseño de vías quimiosintéticas artificiales optimizadas para aplicaciones específicas.
Potenciales avances tecnológicos
- Bioelectroquímica: desarrollo de celdas de combustible microbianas basadas en bacterias quimiosintéticas.
- Captura de carbono: empleo de microorganismos quimioautótrofos para secuestrar CO₂ atmosférico.
- Biología espacial: diseño de sistemas de soporte vital para misiones espaciales basados en ciclos biogeoquímicos impulsados por quimiosíntesis.
- Nanotecnología híbrida: integración de procesos quimiosintéticos con materiales sintéticos para aplicaciones catalíticas.
Cambio climático e importancia creciente
El estudio de la quimiosíntesis adquiere relevancia adicional en el contexto del cambio climático global:
- Las bacterias metanótrofas podrían mitigar emisiones de metano desde humedales, permafrost y océanos.
- Los ecosistemas quimiosintéticos profundos podrían servir como refugios para especies frente al calentamiento superficial.
- La acidificación oceánica podría alterar el funcionamiento de simbiosis quimiosintéticas marinas.
Conclusión
La quimiosíntesis representa una de las adaptaciones metabólicas más extraordinarias de la vida en la Tierra. Desde su descubrimiento por Sergei Winogradsky en el siglo XIX hasta las investigaciones actuales en fumarolas hidrotermales y cuevas aisladas, este proceso ha transformado radicalmente nuestra comprensión sobre los límites de la vida y la diversidad de estrategias que los organismos han desarrollado para obtener energía.
A diferencia de la fotosíntesis, que depende completamente de la luz solar, la quimiosíntesis permite a ciertos microorganismos prosperar en la oscuridad absoluta, utilizando la energía química almacenada en compuestos inorgánicos reducidos. Esta independencia de la luz solar no solo ha permitido colonizar ambientes extremos como el fondo oceánico profundo, sino que probablemente representó el primer tipo de metabolismo autótrofo en los orígenes de la vida hace más de 3.800 millones de años.
Los organismos quimiosintéticos desempeñan roles ecológicos cruciales en el ciclado de nutrientes, la regulación climática y el sostenimiento de ecosistemas únicos. Su estudio continúa revelando aplicaciones biotecnológicas en campos tan diversos como la biorremediación, el tratamiento de aguas, la minería sustentable y la producción de biocombustibles. Además, la existencia de metabolismos quimiosintéticos ha ampliado dramáticamente nuestra búsqueda de vida extraterrestre, sugiriendo que océanos subsuperficiales en lunas heladas podrían albergar comunidades microbianas similares a las de nuestras fumarolas hidrotermales.
Para estudiantes de bachillerato, comprender la quimiosíntesis no solo enriquece el conocimiento sobre metabolismo celular y ciclos biogeoquímicos, sino que ilustra perfectamente cómo el método científico continúa expandiendo los límites de lo que consideramos posible en biología. La historia de la quimiosíntesis nos recuerda que la naturaleza es invariablemente más diversa, adaptable y sorprendente de lo que podemos imaginar.
Reflexión educativa
Como profesora de Biología en educación secundaria, considero que la quimiosíntesis representa uno de los temas más fascinantes pero, paradójicamente, más desatendidos en el currículum educativo actual. Durante mis años de experiencia docente, he observado cómo el estudio de este proceso metabólico abre horizontes completamente nuevos en la mente del alumnado.
Cuando presentamos la fotosíntesis como único proceso de producción primaria, estamos ofreciendo una visión incompleta y terriblemente antropocéntrica de la biosfera. La quimiosíntesis nos recuerda que la vida encuentra caminos insospechados para prosperar y que nuestro planeta alberga ecosistemas extraordinarios que funcionan según principios radicalmente diferentes a los que nos resultan familiares.
He comprobado que introducir la quimiosíntesis en el aula, incluso de forma simplificada, despierta en los estudiantes un renovado interés por la microbiología y la ecología. Les fascina descubrir que existen comunidades biológicas completas en las profundidades oceánicas que nunca han «visto» la luz del sol, o que hay bacterias capaces de «comer piedras» (como describen coloquialmente la oxidación de compuestos inorgánicos).
Este tema ofrece además oportunidades extraordinarias para trabajar la competencia científica de manera interdisciplinar. Conecta con la geología (ciclos biogeoquímicos, tectónica de placas), la química (reacciones redox, termoenergética), la física (gradientes energéticos) y hasta con la astronomía (astrobiología). También permite abordar cuestiones filosóficas sobre los límites y definición de la vida o reflexionar sobre nuestra posición en el universo.
En mis clases, intento complementar las explicaciones teóricas con prácticas sencillas, como el cultivo de bacterias nitrificantes del suelo o la observación de biofilms de bacterias oxidantes del hierro en arroyos cercanos al instituto. Estas experiencias «tangibles» ayudan enormemente a que el alumnado interiorice estos conceptos abstractos.
Me atrevería a decir que, en un mundo donde enfrentamos retos ecológicos sin precedentes, necesitamos estudiantes capaces de comprender la extraordinaria diversidad de estrategias metabólicas que la evolución ha desarrollado. Quién sabe si las soluciones a algunos de nuestros problemas actuales no vendrán precisamente de la comprensión y aplicación de estos procesos quimiosintéticos, marginales pero fundamentales para el funcionamiento global de nuestro planeta.
Por ello, animo a todos mis colegas docentes a profundizar en este apasionante tema y a introducirlo en sus aulas con el entusiasmo que merece. La quimiosíntesis nos recuerda que la vida es aún más extraordinaria, resiliente y diversa de lo que habitualmente enseñamos.
Referencias bibliográficas
Anantharaman, K., Breier, J. A., & Dick, G. J. (2016). Metagenomic resolution of microbial functions in deep-sea hydrothermal plumes across the Eastern Lau Spreading Center. The ISME Journal, 10(1), 225-239.
Cavanaugh, C. M., McKiness, Z. P., Newton, I. L., & Stewart, F. J. (2006). Marine chemosynthetic symbioses. The Prokaryotes, 1, 475-507.
Childress, J. J., & Girguis, P. R. (2011). The metabolic demands of endosymbiotic chemoautotrophic metabolism on host physiological capacities. Journal of Experimental Biology, 214(2), 312-325.
Dick, G. J. (2019). The microbiomes of deep-sea hydrothermal vents: distributed globally, shaped locally. Nature Reviews Microbiology, 17(5), 271-283.
Dubilier, N., Bergin, C., & Lott, C. (2008). Symbiotic diversity in marine animals: the art of harnessing chemosynthesis. Nature Reviews Microbiology, 6(10), 725-740.
Hügler, M., & Sievert, S. M. (2011). Beyond the Calvin cycle: autotrophic carbon fixation in the ocean. Annual Review of Marine Science, 3, 261-289.
Jannasch, H. W. (1985). The chemosynthetic support of life and the microbial diversity at deep-sea hydrothermal vents. Proceedings of the Royal Society of London B, 225(1240), 277-297.
Jørgensen, B. B., & Boetius, A. (2007). Feast and famine—microbial life in the deep-sea bed. Nature Reviews Microbiology, 5(10), 770-781.
Knittel, K., & Boetius, A. (2009). Anaerobic oxidation of methane: progress with an unknown process. Annual Review of Microbiology, 63, 311-334.
Levin, L. A. (2005). Ecology of cold seep sediments: interactions of fauna with flow, chemistry and microbes. Oceanography and Marine Biology: An Annual Review, 43, 1-46.
Martin, W., Baross, J., Kelley, D., & Russell, M. J. (2008). Hydrothermal vents and the origin of life. Nature Reviews Microbiology, 6(11), 805-814.
Nakagawa, S., & Takai, K. (2008). Deep-sea vent chemoautotrophs: diversity, biochemistry and ecological significance. FEMS Microbiology Ecology, 65(1), 1-14.
Orcutt, B. N., Sylvan, J. B., Knab, N. J., & Edwards, K. J. (2011). Microbial ecology of the dark ocean above, at, and below the seafloor. Microbiology and Molecular Biology Reviews, 75(2), 361-422.
Orphan, V. J., House, C. H., Hinrichs, K. U., McKeegan, K. D., & DeLong, E. F. (2002). Multiple archaeal groups mediate methane oxidation in anoxic cold seep sediments. Proceedings of the National Academy of Sciences, 99(11), 7663-7668.
Petersen, J. M., Zielinski, F. U., Pape, T., Seifert, R., Moraru, C., Amann, R., Hourdez, S., Girguis, P. R., Wankel, S. D., Barbe, V., Pelletier, E., Fink, D., Borowski, C., Bach, W., & Dubilier, N. (2011). Hydrogen is an energy source for hydrothermal vent symbioses. Nature, 476(7359), 176-180.
Russell, M. J., & Martin, W. (2004). The rocky roots of the acetyl-CoA pathway. Trends in Biochemical Sciences, 29(7), 358-363.
Schrenk, M. O., Brazelton, W. J., & Lang, S. Q. (2013). Serpentinization, carbon, and deep life. Reviews in Mineralogy and Geochemistry, 75(1), 575-606.
Takai, K., Nakamura, K., Toki, T., Tsunogai, U., Miyazaki, M., Miyazaki, J., Hirayama, H., Nakagawa, S., Nunoura, T., & Horikoshi, K. (2008). Cell proliferation at 122°C and isotopically heavy CH₄ production by a hyperthermophilic methanogen under high-pressure cultivation. Proceedings of the National Academy of Sciences, 105(31), 10949-10954.
Van Dover, C. L. (2000). The Ecology of Deep-Sea Hydrothermal Vents. Princeton University Press.
Wächtershäuser, G. (2007). On the chemistry and evolution of the pioneer organism. Chemistry & Biodiversity, 4(4), 584-602.
