
Los sauropodomorfos constituyen uno de los grupos más fascinantes y exitosos de dinosaurios que jamás hayan pisado nuestro planeta. Estos magníficos herbívoros, pertenecientes al clado Sauropodomorpha, representan una extraordinaria historia evolutiva que abarca desde pequeños bípedos del Triásico tardío hasta los titanes más grandes que han caminado sobre la superficie terrestre.
Datos clave sobre sauropodomorfos
- Período de existencia: 231-66 millones de años (165 millones de años de historia).
- Longitud máxima registrada: ~35-40 metros (Argentinosaurus, Patagotitan).
- Peso máximo estimado: 70-100 toneladas (titanosaurios gigantes).
- Longitud récord de cuello: Hasta 15 metros (Mamenchisaurus).
- Distribución geográfica: Todos los continentes excepto Antártida (aunque hay indicios).
- Número de géneros conocidos: Más de 300 géneros descritos.
- Consumo diario de alimento: 200-400 kg de vegetación para adultos grandes.
Su nombre, derivado del griego, significa literalmente «forma de pie de lagarto», aunque su legado trasciende enormemente esta simple descripción anatómica.
Los sauropodomorfos representan uno de los grupos más exitosos dentro de los dinosaurios, dominando los ecosistemas herbívoros durante millones de años.
¿Qué son los sauropodomorfos?
Los sauropodomorfos son un clado de dinosaurios herbívoros del orden Saurischia que vivieron desde el Triásico Superior (hace 231 millones de años) hasta el final del Cretácico. Se caracterizan por sus cuellos elongados, cabezas pequeñas y la evolución hacia el gigantismo extremo, incluyendo los vertebrados terrestres más grandes conocidos, como los saurópodos.
La historia de los sauropodomorfos es, en muchos sentidos, la historia de una adaptación evolutiva sin precedentes. Estos dinosaurios iniciaron su camino evolutivo como criaturas relativamente modestas en tamaño, pero eventualmente desarrollaron características que les permitirían convertirse en los vertebrados terrestres más grandes conocidos. Sus cuellos elongados, que en algunas especies podían extenderse hasta 15 metros de longitud, les otorgaban acceso a vegetación inaccesible para otros herbívoros, mientras que sus cuerpos masivos les concedían una defensa natural contra depredadores.
Los sauropodomorfos no solo son notables por su tamaño, sino por su increible diversidad y su amplia distribución geográfica. Durante más de 150 millones de años, estos dinosaurios prosperaron en prácticamente todos los continentes, adaptándose a diversos ecosistemas y condiciones climáticas. Su extición, junto con la mayoría de los dinosaurios no avianos, ocurrió hace aproximadamenete 66 millones de años durante el evento de extinción masiva del Cretácico-Paleógeno.
En este artículo, exploraremos en profundidad el fascinante mundo de los sauropodomorfos, desde su clasificación taxonómica y características anatómicas, hasta su comportamiento, evolución y el impacto que han tenido en nuestra comprensión de la historia natural. Acompáñanos en este viaje a través del tiempo para descubrir los secretos de estos gigantes prehistóricos que una vez dominaron los ecosistemas terrestres de nuestro planeta.
Clasificación taxonómica: ubicando a los sauropodomorfos en el árbol de la vida
La clasificación de los sauropodomorfos dentro del complejo árbol filogenético de los dinosaurios ha sido objeto de numerosas revisiones a lo largo de las últimas décadas, reflejando el dinamismo propio de la paleontología como ciencia. Actualmente, y de acuerdo con los consensos científicos más recientes, podemos establecer la siguiente clasificación taxonómica:
- Reino: Animalia.
- Filo: Chordata.
- Clase: Sauropsida.
- Superorden: Dinosauria.
- Orden: Saurischia.
- Suborden: Sauropodomorpha.
El clado Sauropodomorpha se divide tradicionalmente en dos grandes grupos:
- Prosaurópodos (o «prosauropodos»): Representan las formas más primitivas y antiguas del linaje. Si bien este término sigue siendo de uso común en contextos divulgativos, los paleontólogos modernos consideran que los «prosaurópodos» constituyen un grupo parafilético, es decir, no incluyen a todos los descendientes de su ancestro común. Entre los representantes más conocidos encontramos géneros como Plateosaurus, Massospondylus y Anchisaurus.
- Saurópodos: Representan las formas más derivadas y especializadas, caracterizadas principalmente por su gran tamaño y cuadripedalismo obligado. Este grupo incluye a los dinosaurios más grandes que han existido, como Diplodocus, Brachiosaurus, Argentinosaurus y Patagotitan.
Es importante señalar que esta división tradicional ha sido cuestionada en estudios recientes, proponiendo clasificaciones alternativas que reflejan mejor las relaciones evolutivas dentro del grupo. La taxonomía actual reconoce diversos clados dentro de Sauropodomorpha, incluyendo Plateosauridae, Massospondylidae, Diplodocoidea y Macronaria, entre otros.
La filogenia de los sauropodomorfos continúa siendo un campo de investigación activo, donde nuevos descubrimientos fósiles y metodologías analíticas más sofisticadas permiten refinar constantemente nuestra comprensión de las relaciones evolutivas entre estos fascinantes dinosaurios.
Tabla comparativa: Prosaurópodos vs. saurópodos
| Característica | Prosaurópodos | Saurópodos |
|---|---|---|
| Período geológico | Triásico Superior – Jurásico Inferior | Jurásico – Cretácico Superior |
| Tamaño promedio | 2-10 metros de longitud | 15-35 metros de longitud |
| Postura | Bípeda facultativa / cuadrúpeda | Exclusivamente cuadrúpeda |
| Longitud del cuello | Moderadamente largo (10-12 vértebras) | Extremadamente largo (hasta 19 vértebras) |
| Dentición | Heterodóntica, dientes con serrado | Homodóntica, dientes tipo clavija o espátula |
| Ejemplos representativos | Plateosaurus, Massospondylus | Diplodocus, Brachiosaurus, Argentinosaurus |
| Diversidad de especies | ~50 géneros conocidos | ~300 géneros conocidos |
Características anatómicas: La estructura de los gigantes
Los sauropodomorfos presentan una serie de características anatómicas distintivas que evolucionaron a lo largo de millones de años, permitiéndoles adaptarse a diversos nichos ecológicos y, en el caso de los saurópodos, alcanzar tamaños gigantescos.
Un aspecto fundamental en la evolución anatómica de los sauropodomorfos fue el desarrollo de proporciones corporales óptimas para soportar masas de hasta 70-100 toneladas. Sus huesos no eran simplemente versiones agrandadas de dinosaurios más pequeños, sino estructuras rediseñadas con cavidades de aire (estructuras pneumáticas) que reducían peso sin comprometer resistencia estructural. Las vértebras presentaban complejos sistemas de arcos y cavidades que las hacían simultáneamente ligeras y robustas, un diseño biomecánico que ha inspirado aplicaciones en ingeniería moderna de estructuras ligeras de alta resistencia.
A continuación, analizaremos los principales rasgos anatómicos que definieron a este grupo:
Cráneo y dentición
El cráneo de los sauropodomorfos era relativamente pequeño en proporción al resto del cuerpo, especialmente en las formas más derivadas. Esta característica se acentuó progresivamente a lo largo de su evolución, culminando en los saurópodos con cráneos extremadamente reducidos en relación con su masa corporal.
La dentición varió considerablemente dentro del grupo:
- Los prosaurópodos primitivos poseían dientes heterodónticos (de diferentes formas), incluyendo algunos ligeramente serrados que sugieren una dieta omnívora en las formas más basales.
- A medida que evolucionaron, desarrollaron dientes en forma de cuchara o espátula, perfectamente adaptados para arrancar materia vegetal.
- Los saurópodos avanzados presentaban dientes en forma de lápiz o clavija, sin capacidad de masticación, indicando que tragaban el alimento entero para ser procesado posteriormente en su sistema digestivo.
Cuello y columna vertebral
Quizás la característica más distintiva de los sauropodomorfos avanzados fue su cuello extremadamente largo. Esta elongación se logró mediante:
- Un aumento en el número de vértebras cervicales (en algunos casos hasta 19, mientras que la mayoría de los vertebrados poseen 7).
- La neumatización de las vértebras, es decir, la presencia de cavidades llenas de aire que reducían el peso sin comprometer la resistencia estructural.
- Un sistema de articulaciones altamente especializado que permitía un considerable rango de movimiento.
Las vértebras dorsales y caudales también experimentaron modificaciones significativas, desarrollando estructuras como las espinas neurales bifurcadas en algunos grupos (como los diplodócidos) o las hipapófisis en otros, que servían como puntos de anclaje para una musculatura cada vez más compleja.
Especies con los cuellos más largos
| Especie | Longitud del Cuello | Vértebras Cervicales | Período/Localización |
|---|---|---|---|
| Mamenchisaurus sinocanadorum | ~15 metros | 18 | Jurásico Superior, China |
| Supersaurus vivianae | ~13-14 metros | 15 | Jurásico Superior, EE.UU. |
| Sauroposeidon proteles | ~11-12 metros | 13-14 | Cretácico Inferior, EE.UU. |
| Diplodocus carnegii | ~8 metros | 15 | Jurásico Superior, EE.UU. |
Extremidades y postura
La evolución de las extremidades en los sauropodomorfos refleja uno de los cambios posturales más significativos en la historia de los vertebrados terrestres:
- Los sauropodomorfos basales eran predominantemente bípedos facultativos, capaces de desplazarse tanto sobre dos como sobre cuatro patas.
- La transición hacia el cuadripedalismo obligado en los saurópodos implicó profundas modificaciones anatómicas, incluyendo la robustecimiento de las extremidades anteriores y la modificación de la estructura pélvica.
- Las extremidades de los saurópodos evolucionaron como columnas prácticamente rectas, optimizadas para soportar el enorme peso corporal.
- Los pies desarrollaron una configuración única con dedos reducidos, rodeados por un «cojín» de tejido blando similar al de los elefantes actuales, que distribuía eficientemente el peso.
Sistema respiratorio y metabolismo
Los estudios de la anatomía interna de los sauropodomorfos, particularmente de los saurópodos, revelan un sistema respiratorio altamente especializado:
- Presencia de sacos aéreos similares a los de las aves modernas, que permitían una respiración unidireccional altamente eficiente.
- Un sistema de neumatización que se extendía por gran parte del esqueleto axial, creando una red de cavidades conectadas con el sistema respiratorio.
Estas adaptaciones sugieren un metabolismo relativamente elevado, especialmente considerando los requerimientos energéticos asociados al gigantismo.
A nivel microscópico, el estudio de las células fosilizadas ha revelado información valiosa sobre el metabolismo de estos gigantes.
Las investigaciones más recientes, basadas en análisis de líneas de crecimiento óseo (similares a los anillos de los árboles) y modelos termométricos, sugieren que los sauropodomorfos grandes probablemente exhibían gigantotermia o mesotermia. Este estado metabólico intermedio implica que, aunque no generaban activamente calor metabólico como los mamíferos (endotermia verdadera), su enorme masa corporal les permitía mantener temperaturas internas estables y elevadas mediante inercia térmica. Un saurópodo de 50 toneladas tardaría días en enfriarse o calentarse significativamente, funcionando efectivamente como un «horno de sangre tibia» que operaba a temperaturas corporales relativamente constantes de 36-38°C, óptimas para procesos fisiológicos eficientes. Esta estrategia termorreguladora habría sido imposible para dinosaurios más pequeños, representando una ventaja evolutiva exclusiva del gigantismo extremo.
Integumento y termorregulación
Tradicionalmente, se ha representado a los sauropodomorfos con una piel escamosa similar a la de los reptiles actuales. Sin embargo, la evidencia fósil directa sobre su integumento es escasa. Algunos investigadores han propuesto que podrían haber poseído estructuras filamentosas primitivas en ciertas partes del cuerpo, al menos en formas basales, aunque esta hipótesis sigue siendo especulativa.
La termorregulación en animales de tal magnitud plantea desafíos únicos. Los estudios sugieren que los saurópodos más grandes probablemente empleaban la inercia térmica como principal mecanismo termorregulador, manteniend una temperatura corporal relativamente constante debido a su enorme masa.
Estas extraordinarias adaptaciones anatómicas permitieron a los sauropodomorfos, especialmente a los saurópodos, explotar nichos ecológicos inaccesibles para otros herbívoros, convirtiéndose en uno de los grupos de vertebrados terrestres más exitosos durante la era Mesozoica.
Evolución y diversificación: El camino hacia el gigantismo
La historia evolutiva de los sauropodomorfos representa uno de los ejemplos más fascinantes de radiación adaptativa en el registro fósil, desde sus modestos orígenes en el Triásico Superior hasta el apogeo del gigantismo en el Jurásico y Cretácico. Este proceso evolutivo no fue lineal ni uniforme, sino una compleja trayectoria marcada por innovaciones anatómicas, cambios ecológicos y eventos de diversificación y extinción.
Los cambios en la estructura de la Tierra durante el Mesozoico, incluyendo la fragmentación de Pangea, influyeron decisivamente en su distribución.
Radiación del Jurásico: La edad de oro de los saurópodos
El Jurásico representa el período de mayor esplendor y diversificación de los sauropodomorfos, particularmente de los saurópodos verdaderos. Durante esta era, que se extiende desde hace 201 hasta 145 millones de años, estos gigantes alcanzaron su máxima expresión evolutiva y colonizaron prácticamente todos los continentes.
Los primeros saurópodos verdaderos, como Vulcanodon y Barapasaurus, aparecieron en el Jurásico Inferior (hace aproximadamente 200-190 millones de años). Estas formas transicionales aún conservaban algunas características de los prosaurópodos, pero ya mostraban la postura exclusivamente cuadrúpeda y el alargamiento cervical característico del grupo.
Durante el Jurásico Medio y Superior se produjo una extraordinaria radiación adaptativa que dio origen a las principales familias de saurópodos. Los Diplodocoidea desarrollaron cuellos extremadamente largos con cabezas pequeñas y colas en forma de látigo, como se observa en géneros icónicos como Diplodocus, Apatosaurus y Brachiosaurus. Por su parte, los Macronaria evolucionaron hacia formas masivas con extremidades columnares robustas, culminando en gigantes como Camarasaurus.
El registro fósil de la Formación Morrison en Norteamérica, con más de 150 millones de años de antigüedad, proporciona una ventana excepcional a este ecosistema dominado por saurópodos. Hasta 13 especies diferentes de sauropodomorfos coexistían en estas regiones, cada una ocupando nichos ecológicos ligeramente diferentes según la altura de ramoneo, el tipo de vegetación consumida y las preferencias de hábitat.
Los factores que impulsaron esta radiación fueron múltiples: niveles atmosféricos de CO₂ más elevados que favorecían una vegetación exuberante, temperaturas cálidas globales sin casquetes polares significativos, y la ausencia de competidores herbívoros de gran tamaño. Los sauropodomorfos del Jurásico literalmente remodelaban los paisajes, actuando como «ingenieros ecosistémicos» al derribar árboles y crear claros que favorecían la diversidad vegetal.
El Cretácico: Diversificación global y especialización regional
El período Cretácico (145-66 millones de años atrás) marcó una fase de transformación profunda en la historia evolutiva de los sauropodomorfos. Aunque los prosaurópodos ya se habían extinguido completamente a principios de este período, los saurópodos experimentaron una segunda gran radiación, particularmente en el hemisferio sur.
Los Titanosauria se convirtieron en el grupo dominante del Cretácico, especialmente en los continentes del antiguo Gondwana (Sudamérica, África, India y Madagascar). Estos sauropodomorfos desarrollaron características únicas, incluyendo osteodermos dérmicos (placas óseas en la piel) que proporcionaban protección adicional contra depredadores. Especies como Argentinosaurus, Dreadnoughtus y Patagotitan se encuentran entre los animales terrestres más grandes jamás documentados, con estimaciones de peso que alcanzan las 70-100 toneladas.
La fragmentación del supercontinente Pangea durante el Cretácico condujo a patrones de endemismo y evolución regional. En Sudamérica, los titanosaurios desarrollaron formas extremadamente diversas, desde especies relativamente pequeñas de 6-8 metros hasta los gigantes absolutos mencionados anteriormente. En Europa, formas insulares como Magyarosaurus experimentaron enanismo insular, reduciendo su tamaño a apenas 6 metros debido a los recursos limitados de sus hábitats isleños.
Un desarrollo evolutivo fascinante del Cretácico fue la aparición de titanosaurios con dentición altamente especializada. Especies como Nigersaurus desarrollaron baterías dentales con más de 500 dientes de reemplazo activo, adaptados para un ramoneo de baja altura similar al de los herbívoros modernos que consumen gramíneas. Esta especialización sugiere que algunos sauropodomorfos del Cretácico ocupaban nichos ecológicos comparables a los de los ungulados actuales.
El final del Cretácico vio una curiosa disminución en la diversidad de sauropodomorfos en Norteamérica y Asia, mientras que en Sudamérica y África estos dinosaurios continuaban siendo abundantes. Esta distribución asimétrica hace que el debate sobre las causas de su extinción final sea particularmente intrigante, sugiriendo que factores regionales además del impacto del asteroide pudieron haber desempeñado un papel importante.
Orígenes triásicos
Los primeros sauropodomorfos aparecieron hace aproximadamente 231-228 millones de años, durante el Triásico Superior (Carniense). Estos primeros representantes, como Saturnalia tupiniquim y Panphagia protos, eran animales relativamente pequeños, con longitudes corporales de 1,5 a 2 metros. Anatómicamente, se caracterizaban por:
- Una postura bípeda predominante.
- Un cráneo pequeño con dientes ligeramente heterodónticos.
- Manos prensiles con un pulgar ligeramente modificado.
- Una talla modesta en comparación con sus descendientes.
Estos rasgos primitivos sugieren que los primeros sauropodomorfos eran omnívoros oportunistas, probablemente capaces de explotar recursos tanto vegetales como animales, ocupando nichos ecológicos intermedios entre los carnívoros y los herbívoros especializados.
Radiación durante el triásico tardío
Durante el Noriense y el Rétiense (etapas finales del Triásico), los sauropodomorfos experimentaron una notable diversificación. Surgieron formas como Plateosaurus, Riojasaurus y Mussaurus, que representaban un paso evolutivo significativo:
- Incremento en el tamaño corporal (algunos superaban los 6-8 metros)
- Mayor especialización hacia una dieta herbívora
- Desarrollo de cuellos más largos, aunque todavía modestos en comparación con los saurópodos posteriores
- Capacidad para adoptar tanto posturas bípedas como cuadrúpedas (bipedalismo facultativo)
Esta radiación coincidió con la proliferación de las gimnospermas (especialmente cicadáceas y coníferas) que constituirían su principal fuente de alimento. Los sauropodomorfos triásicos lograron extenderse ampliamente, con fósiles documentados en lo que hoy es Europa, América del Sur, África, Asia y la Antártida, demostrando su éxito evolutivo temprano.
La transición prosaurópodo-saurópodo
El límite Triásico-Jurásico (hace aproximadamente 201 millones de años) marcó un punto de inflexión en la historia de los vertebrados terrestres, con la extinción de numerosos grupos de arcosaurios no dinosaurianos. Los sauropodomorfos, sin embargo, sobrevivieron a este evento de extinción y experimentaron una importante transición evolutiva durante el Jurásico Inferior.
Formas transicionales como Anchisaurus, Massospondylus y especialmente Melanorosaurus y Lessemsaurus muestran características intermedias entre los prosaurópodos clásicos y los verdaderos saurópodos:
- Incremento en la robustez de las extremidades anteriores.
- Modificaciones en la estructura pélvica.
- Mayor neumatización del esqueleto axial.
- Tendencia hacia el cuadripedalismo obligado.
La aparición de los verdaderos saurópodos
Los primeros saurópodos propiamente dichos aparecieron en el Jurásico Inferior, con géneros como Vulcanodon y Barapasaurus, que ya exhibían las características distintivas del grupo:
- Cuadripedalismo obligado
- Cuello significativamente elongado
- Cráneo relativamente pequeño
- Extremidades columnares
- Mayor neumatización del esqueleto para reducir el peso
- Dentición especializada en forma de clavija o lápiz
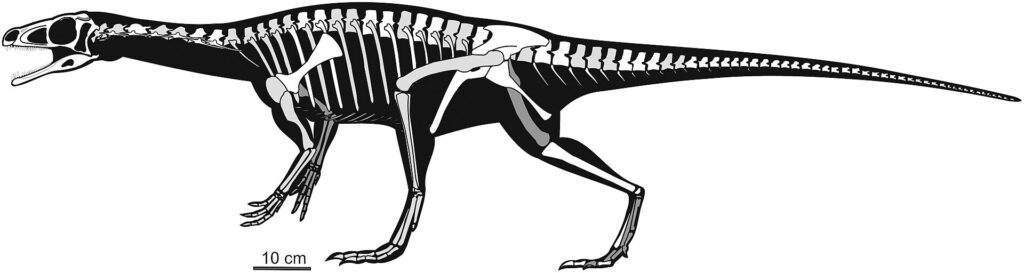
El registro fósil muestra que la transición hacia el verdadero gigantismo ocurrió con notable rapidez en términos evolutivos. Para el Jurásico Medio (hace aproximadamente 170-165 millones de años), ya existían saurópodos de gran tamaño como Mamenchisaurus en Asia y los primeros representantes de linajes que dominarían el Jurásico Tardío.
La edad de oro de los saurópodos
El Jurásico Superior (hace 163-145 millones de años) representa el apogeo en la diversificación de los saurópodos, con la aparición de los principales linajes que definirían al grupo:
- Diplodócidos: Caracterizados por cuellos y colas extremadamente largos, cráneos alargados con dientes concentrados en la parte frontal de la mandíbula. Ejemplos notables incluyen Diplodocus, Apatosaurus y Barosaurus.
- Braquiosáuridos: Distinguidos por sus extremidades anteriores más largas que las posteriores, generando una postura inclinada, y cuellos extremadamente altos. El género tipo Brachiosaurus y el famoso Giraffatitan son representantes emblemáticos.
- Camarasáuridos: Con cráneos más robustos y cuellos relativamente más cortos que otros saurópodos contemporáneos. Camarasaurus es el género más conocido de este grupo.
Esta diversificación morfológica y ecológica permitió la coexistencia de diferentes tipos de saurópodos en los mismos ecosistemas, explotando distintos recursos vegetales a diferentes alturas, un fenómeno conocido como partición de nichos ecológicos.
Saurópodos del Cretácico: Diversificación final
Contrariamente a lo que se pensaba en el siglo pasado, los saurópodos no experimentaron un declive durante el Cretácico, sino una nueva radiación adaptativa, especialmente en los continentes del hemisferio sur. El Cretácico (145-66 millones de años) vio la aparición de nuevos grupos especializados:
- Titanosaurios: El grupo más diverso de saurópodos cretácicos, con una extraordinaria distribución global. Incluye a algunos de los dinosaurios más grandes conocidos, como Argentinosaurus, Patagotitan y Puertasaurus. Desarrollaron características únicas como la armadura dérmica (osteodermos) en algunas especies.
- Rebaquisáuridos: Saurópodos especializado con cráneos alargados similares a los de los diplodócidos, pero pertenecientes a un linaje diferente. Rebbachisaurus y Nigersaurus (con su extraordinario aparato bucal adaptado para «pastar») son ejemplos representativos.
- Mamenchisáuridos: Aunque principalmente jurásicos, algunos representantes sobrevivieron hasta el Cretácico Inferior, principalmente en Asia. Se caracterizaban por poseer los cuellos proporcionalmente más largos entre todos los saurópodos.
El camino evolutivo hacia el gigantismo
El gigantismo extremo de los saurópodos plantea fascinantes preguntas evolutivas. Los estudios sugieren que esta tendencia fue posibilitada por una combinación de factores:
- Ventajas ecológicas: El gran tamaño proporcionaba acceso a recursos alimenticios inaccesibles para otros herbívoros y defensa contra depredadores.
- Innovaciones anatómicas: La neumatización del esqueleto, el sistema de sacos aéreos tipo aviano, y la reducción del cráneo disminuían el peso sin comprometer la integridad estructural.
- Fisiología eficiente: Un sistema digestivo altamente especializado que permitía procesar grandes cantidades de vegetación de baja calidad nutritiva.
- Factores ambientales: Los niveles elevados de oxígeno atmosférico y la abundancia de vegetación de alto crecimiento durante el Mesozoico proporcionaron condiciones favorables.
Este proceso evolutivo no estuvo exento de limitaciones físicas y fisiológicas. Existe evidencia de que los saurópodos más grandes operaban cerca de los límites biomecánicos impuestos por las leyes de la física, particularmente en relación con la resistencia ósea y los requerimientos energéticos.
Ecología y comportamiento: La vida de los gigantes
Sauropodomorfos en la península ibérica
España posee uno de los registros fósiles de sauropodomorfos más ricos y diversos de Europa, con yacimientos que abarcan desde el Jurásico hasta el Cretácico Superior. Este patrimonio paleontológico proporciona información valiosa sobre la biogeografía y evolución de estos dinosaurios en el contexto europeo.
Yacimientos del Jurásico
El yacimiento de Peñarroya de Tastavins (Teruel) ha proporcionado restos de saurópodos del Jurásico Superior, incluyendo vértebras y elementos apendiculares que sugieren la presencia de formas relacionadas con diplodócidos. En la Formación de Villar del Arzobispo (Valencia y Cuenca) se han documentado abundantes icnitas (huellas) de sauropodomorfos que revelan comportamientos de manada y rutas migratorias.
La Sierra de los Cameros (La Rioja) conserva uno de los registros de icnitas más espectaculares, con rastrilladas que muestran el paso de manadas completas de sauropodomorfos, incluyendo individuos de diferentes edades y tamaños caminando sincronizadamente.
Sauropodomorfos del Cretácico ibérico
El Cretácico Inferior de la Formación de El Castellar (Teruel) ha proporcionado restos de saurópodos titanosauriformes, representando las formas de transición entre saurópodos más primitivos y los titanosaurios verdaderos que dominarían el Cretácico Superior.
Los yacimientos del Cretácico Superior en Cataluña (cuenca del Tremp) son particularmente importantes, habiendo proporcionado restos de titanosaurios que vivieron en los últimos millones de años antes de la extinción. Especies como Lirainosaurus astibiae del País Vasco representan formas de tamaño medio (6-7 metros) que habitaban ecosistemas costeros y deltaicos.
Importancia científica y cultural
El estudio de los sauropodomorfos ibéricos ha contribuido significativamente al conocimiento de la biogeografía europea durante el Mesozoico. La posición de Iberia como puente entre África y Europa permitió intercambios faunísticos que se reflejan en la mezcla de linajes gondwánicos y laurásicos presentes en el registro fósil español.
Numerosos museos españoles exhiben réplicas y restos originales de estos dinosaurios, incluyendo el Museo Jurásico de Asturias, Dinópolis en Teruel, y el Museo de Ciencias Naturales de Madrid, facilitando la divulgación científica y conectando al público general con este fascinante patrimonio paleontológico nacional.
Extinción y legado: El Final de una era
El evento K-Pg y la desaparición de los sauropodomorfos
Hace aproximadamente 66 millones de años, el impacto de un asteroide de unos 10 kilómetros de diámetro en la península de Yucatán (México) desencadenó el evento de extinción masiva del Cretácico-Paleógeno (K-Pg), también conocido como extinción K-T. Este cataclismo marcó el final de los sauropodomorfos junto con la mayoría de los dinosaurios no avianos.
Los efectos inmediatos del impacto incluyeron:
- Invierno de impacto: Partículas en suspensión bloquearon la luz solar durante meses o años, colapsando las cadenas tróficas basadas en fotosíntesis
- Incendios globales: Ondas de calor radiante iniciaron incendios forestales en múltiples continentes
- Lluvia ácida: Vaporizó rocas que contenían azufre y modificaron la química oceánica y atmosférica
- Tsunamis masivos: Olas gigantes devastaron regiones costeras donde vivían muchos titanosaurios
Para los sauropodomorfos, dependientes de enormes cantidades de vegetación diaria (se estima que un saurópodo de 50 toneladas necesitaba consumir 200-400 kg de plantas diariamente), el colapso de la productividad vegetal fue especialmente catastrófico. Su gran tamaño, que había sido una ventaja evolutiva durante 150 millones de años, se convirtió en una desventaja letal cuando la base de la cadena alimentaria desapareció.
Supervivientes improbables
Curiosamente, estudios recientes sugieren que algunos linajes de sauropodomorfos podrían haber sobrevivido brevemente después del impacto inicial. Restos fósiles en la Formación Scollard de Alberta (Canadá) y en la Formación Ojo Alamo de Nuevo México (EE.UU.) han sido datados tentativamemente en el Paleoceno temprano, sugiriendo supervivencias locales de quizás miles de años después del evento K-Pg.
Sin embargo, estas afirmaciones son controvertidas y la mayoría de paleontólogos consideran que estos fósiles fueron redepositados desde estratos más antiguos. El consenso científico actual sostiene que los sauropodomorfos se extinguieron completamente en el límite K-Pg, sin dejar descendientes supervivientes.
Legado científico y cultural
El legado de los sauropodomorfos trasciende su extinción física. Han contribuido enormemente a nuestro entendimiento de:
- Biomecánica a gran escala: Cómo funcionan los cuerpos extremadamente grandes y los límites físicos del gigantismo terrestre.
- Evolución adaptativa: Procesos de especialización y radiación en respuesta a oportunidades ecológicas.
- Paleoecología: Reconstrucción de ecosistemas antiguos y redes tróficas del Mesozoico.
- Biogeografía: Distribución continental de fauna y efectos de la deriva continental.
Culturalmente, los sauropodomorfos capturan la imaginación pública como pocos otros grupos de organismos extintos. Desde las primeras reconstrucciones victorianas hasta las películas y documentales modernos, estos gigantes prehistóricos simbolizan la inmensidad del tiempo geológico y la extraordinaria diversidad de la vida en la Tierra.
En el contexto educativo español, el estudio de los sauropodomorfos forma parte integral de los currículos de biología, geología y ciencias naturales en secundaria y bachillerato, proporcionando ejemplos tangibles de evolución, adaptación y los procesos que dan forma a la biodiversidad.
Estrategias de alimentación y dieta
Los sauropodomorfos fueron herbívoros especializados cuyas estrategias alimentarias evolucionaron en paralelo con sus cambios anatómicos. Su dieta consistía principalmente en gimnospermas (coníferas, cycadas, ginkgos) y helechos, la vegetación dominante durante el Mesozoico. Las angiospermas (plantas con flores) solo aparecieron en el Cretácico, y algunos sauropodomorfos tardíos las incorporaron a su dieta.
El procesamiento del alimento variaba significativamente entre grupos. Los prosaurópodos, con su dentición heterodóntica, podían realizar cierta masticación preliminar. En contraste, los saurópodos empleaban una estrategia de «ramoneo sin masticación»: sus dientes tipo clavija o espátula servían únicamente para arrancar vegetación, que era tragada entera y procesada en un sistema digestivo fermentativo extremadamente largo.
Los gastrolitos (piedras gastrales) jugaban un papel fundamental en la digestión. Estos guijarros redondeados y pulidos por el desgaste se han encontrado asociados a esqueletos de sauropodomorfos, confirmando su función de moler mecánicamente la materia vegetal en el estómago muscular, de manera similar a las aves modernas.
Estudios biomecánicos sugieren diferentes estrategias de ramoneo según la especie. Los diplodócidos, con cuellos extremadamente largos y flexibles, probablemente ramoneaban horizontalmente en amplias áreas, mientras que los braquiosáuridos, con extremidades anteriores más largas que las posteriores, estaban adaptados para alimentarse de vegetación alta, alcanzando copas de árboles a 9-10 metros del suelo.
Comportamiento social y reproducción
La evidencia paleontológica sugiere que muchos sauropodomorfos eran animales gregarios que vivían en grupos o manadas. Pistas fósiles (icnitas) preservadas muestran caminos de múltiples individuos caminando en la misma dirección, con animales juveniles protegidos en el centro del grupo, un patrón de comportamiento similar al de elefantes y otros grandes herbívoros actuales.
Los nidos y huevos fósiles de sauropodomorfos, particularmente de titanosaurios, revelan estrategias reproductivas fascinantes. Ponían huevos relativamente pequeños (del tamaño de un balón de fútbol) en nidos comunales, con múltiples hembras depositando sus huevos en el mismo sitio. Las crías eclosionaban con apenas 3-5 kg de peso, experimentando después un crecimiento explosivo que podía añadir más de 2 toneladas anuales durante la juventud.
El cuidado parental directo era probablemente limitado o inexistente en la mayoría de especies, dado el riesgo que supondría para las crías ser pisoteadas accidentalmente por adultos de 30-50 toneladas. En cambio, se ha propuesto que los juveniles formaban grupos separados en áreas de vegetación densa que proporcionaban protección natural.
Depredadores y estrategias defensivas
A pesar de su tamaño masivo, los sauropodomorfos enfrentaban amenazas de grandes terópodos depredadores. Mientras que los adultos completamente desarrollados tenían pocos enemigos naturales debido a su tamaño, los juveniles y subadultos eran vulnerables a ataques de carnívoros como los abelisáuridos y carcharodontosáuridos.
Las principales defensas de los sauropodomorfos incluían:
- Tamaño corporal: Un saurópodo adulto de 40-70 toneladas era prácticamente invulnerable a cualquier depredador.
- Comportamiento gregario: Vivir en manadas proporcionaba protección colectiva, especialmente a los jóvenes.
- Cola como arma: Algunos grupos, como los diplodócidos, poseían colas extremadamente largas con estructuras terminales que funcionaban como látigos capaces de generar chasquidos supersónicos.
- Osteodermos: Los titanosaurios del Cretácico desarrollaron placas óseas dérmicas que proporcionaban armadura adicional.
Evidencia de lesiones en huesos fósiles, incluyendo marcas de dientes y fracturas sanadas, confirma que algunos sauropodomorfos sobrevivieron a ataques de depredadores, demostrando su resistencia y capacidad de recuperación.
El éxito evolutivo de los sauropodomorfos, culminando en el gigantismo extremo de los saurópodos, representa uno de los más extraordinarios ejemplos de adaptación en la historia de la vida terrestre, demostrando la capacidad de la selección natural para producir formas que explotan eficientemente los recursos disponibles en su entorno.
Paleobiología y comportamiento: La vida de los gigantes
Reconstruir el comportamiento y la biología de animales extintos hace millones de años constituye uno de los mayores desafíos de la paleontología. Sin embargo, a través del estudio minucioso de la anatomía, tafonomía, huellas fósiles y comparaciones con animales actuales, los científicos han podido inferir aspectos fascinantes sobre la paleobiología de los sauropodomorfos.
Alimentación y digestión
La dieta de los sauropodomorfos experimentó una progresiva especialización hacia el herbivorismo a lo largo de su evolución:
- Los sauropodomorfos basales probablemente eran omnívoros oportunistas, como sugiere su dentición parcialmente heterodóntica.
- Los «prosaurópodos» avanzados ya mostraban adaptaciones claras para una dieta principalmente herbívora, con dientes en forma de espátula adaptados para cortar material vegetal.
- Los saurópodos desarrollaron un sistema de alimentación altamente especializado basado en:
- Arrancado no selectivo de vegetación mediante dientes en forma de clavija.
- Ausencia de masticación, compensada por un extenso procesamiento gástrico.
- Probable presencia de gastrolitos (piedras estomacales) en algunas especies para ayudar en la trituración del alimento.
- Un tracto digestivo extremadamente largo con fermentación bacteriana para procesar celulosa.
Los estudios de coprolitos (excrementos fosilizados) y contenidos estomacales preservados sugieren que los principales componentes de su dieta incluían coníferas, ginkgos, helechos, equisetos y cicadales. Análisis de isótopos estables en dientes y huesos de saurópodos han aportado evidencia adicional sobre sus preferencias alimentarias y hábitos de forrajeo.
Crecimiento y desarrollo
El estudio histológico de los huesos de sauropodomorfos ha revelado patrones de crecimiento sorprendentes:
- A diferencia de los reptiles modernos, los sauropodomorfos experimentaban un crecimiento rápido durante sus primeras etapas de vida, similar al observado en mamíferos y aves.
- Los saurópodos alcanzaban la madurez sexual antes de la madurez esquelética, lo que les permitía reproducirse mientras aún crecían.
- El crecimiento se ralentizaba progresivamente, pero nunca cesaba por completo en algunos taxones, un patrón conocido como crecimiento indeterminado.
- La tasa metabólica necesaria para sostener este rápido crecimiento sugiere un metabolismo significativamente más elevado que el de los reptiles ectotérmicos actuales.
Los juveniles de sauropodomorfo probablemente ocupaban nichos ecológicos diferentes a los adultos, con posturas más bípedas y dietas potencialmente distintas, reduciendo así la competencia intergeneracional.
Reproducción y cuidado parental
Los descubrimientos de huevos, nidos y embriones fósiles han permitido reconstruir aspectos clave de la biología reproductiva de los sauropodomorfos:
- Ponían huevos relativamente pequeños (15-25 cm de diámetro) en relación con su tamaño corporal, una limitación biomecánica que compensaban mediante estrategias de puesta múltiple.
- Los hallazgos en sitios como Auca Mahuevo (Argentina) y Coll de Nargó (España) revelan comportamientos de nidificación comunal, con numerosos nidos agrupados en áreas específicas.
- La estructura porosa de las cáscaras sugiere que los huevos eran enterrados en sustrato, similar a lo observado en cocodrilos y tortugas actuales.
- Existe debate sobre el grado de cuidado parental. Algunos investigadores sugieren un modelo similar al de los cocodrilos modernos, con cierta protección del nido, mientras que otros proponen una estrategia más «r-seleccionada» con escaso o nulo cuidado tras la puesta.
La extrema diferencia de tamaño entre crías y adultos (hasta 2.500 veces en los saurópodos más grandes) plantea interesantes cuestiones sobre la dinámica social y las estrategias de supervivencia juvenil.
Comportamiento Social y Gregario
La evidencia de comportamiento gregario en sauropodomorfos proviene de múltiples fuentes:
- Yacimientos monoespcíficos que contienen múltiples individuos, como los hallados para Plateosaurus en Europa o Mussaurus en Argentina.
- Rastros de huellas paralelas dejadas por varios individuos moviéndose en la misma dirección y al mismo tiempo.
- Evidencia de dimorfismo sexual en algunos taxones, que podría indicar sistemas de apareamiento complejos.
Los estudios sugieren que muchos sauropodomorfos formaban manadas o agregaciones sociales, probablemente estructuradas por edad, como mecanismo de protección contra depredadores y posiblemente para optimizar la explotación de recursos. Sin embargo, el grado de complejidad social variaría entre diferentes taxones y probablemente cambiaría a lo largo del desarrollo ontogenético.
Locomoción y desplazamiento
La locomoción de los sauropodomorfos ha sido objeto de intenso estudio:
- Los sauropodomorfos basales eran predominantemente bípedos, pero capaces de adoptar posturas cuadrúpedas para actividades específicas como alimentarse.
- La transición al cuadripedalismo obligado en saurópodos implicó profundas modificaciones anatómicas, incluyendo la rotación de la mano y cambios en la musculatura asociada.
- Los análisis biomecánicos sugieren velocidades máximas de aproximadamente 20-25 km/h para los saurópodos más ágiles, considerablemente menor que la de terópodos contemporáneos.
- Estudios de huellas fósiles indican que los saurópodos se desplazaban principalmente con un paso amblador, similar al de los elefantes modernos, donde las extremidades del mismo lado se mueven casi sincronizadamente.
Las reconstrucciones de la musculatura axial y apendicular, basadas en marcas de inserción muscular y comparaciones con reptiles y aves modernas, han permitido crear modelos biomecánicos cada vez más precisos sobre cómo se movían estos gigantes.
Termorregulación y fisiología
La fisiología de los sauropodomorfos, particularmente de los saurópodos gigantes, sigue siendo un tema de debate científico:
- El gran tamaño corporal habría proporcionado inercia térmica (gigantotermia), permitiendo mantener una temperatura corporal relativamente estable a pesar de las fluctuaciones ambientales.
- La presencia de un sistema respiratorio similar al de las aves, con sacos aéreos y ventilación unidireccional, sugiere un metabolismo elevado.
- Los análisis histológicos revelan patrones de crecimiento rápido que requerirían tasas metabólicas superiores a las de los reptiles ectotérmicos actuales.
- Estudios de paleotemperatura basados en isótopos de oxígeno en dientes y huesos sugieren temperaturas corporales elevadas y relativamente constantes.
El consenso actual se inclina hacia un modelo fisiológico intermedio: ni completamente ectotérmicos como los reptiles actuales, ni endotérmicos como las aves y mamíferos modernos, sino poseedores de una fisiología mesotérmica o taquimetabólica con características únicas adaptadas a su enorme tamaño.
La paleobiología de los sauropodomorfos continúa siendo un campo dinámico donde nuevos descubrimientos y metodologías analíticas refinadas permiten profundizar nuestra comprensión de cómo vivían estos extraordinarios animales, desafiando constantemente nuestras preconcepciones sobre las limitaciones biológicas y ecológicas.
Distribución geográfica y temporal: Un éxito global
Los sauropodomorfos representan una de las historias de éxito evolutivo más notables en la historia de los vertebrados terrestres, no solo por su longevidad temporal sino también por su extraordinaria distribución geográfica. A lo largo de más de 150 millones de años de evolución, estos dinosaurios conquistaron prácticamente todos los continentes, adaptándose a una amplia variedad de ecosistemas y condiciones paleoambientales.
Distribución temporal
La historia temporal de los sauropodomorfos puede dividirse en varias fases distintivas:
Fase inicial (Triásico Superior: 231-201 millones de años)
Los primeros sauropodomorfos aparecen en el registro fósil durante el Carniense tardío, con formas basales como Saturnalia y Panphagia en lo que hoy es Sudamérica. Durante el Noriense y Rétiense, experimentaron una notable radiación adaptativa, diversificándose en numerosos géneros como Plateosaurus, Unaysaurus, Riojasaurus y Coloradisaurus, entre otros.
Esta primera fase culminó con el evento de extinción del Triásico-Jurásico, que aunque eliminó a numerosos grupos de arcosaurios, parece haber afectado relativamente poco a los sauropodomorfos, permitiéndoles diversificarse aún más en el período siguiente.
Fase de transición (Jurásico Inferior: 201-174 millones de años)
El Jurásico Inferior representa un período crucial en la evolución de los sauropodomorfos, marcado por la transición gradual desde las formas basales («prosaurópodos») hacia los verdaderos saurópodos. Durante esta época encontramos una mezcla de:
- Sauropodomorfos basales persistentes como Massospondylus y Anchisaurus.
- Formas transicionales como Melanorosaurus y Lessemsaurus.
- Los primeros saurópodos verdaderos como Vulcanodon y Barapasaurus.
Esta fase representa la consolidación de las innovaciones anatómicas que permitirían el posterior gigantismo del grupo.
Fase de diversificación (Jurásico Medio y Superior: 174-145 millones de años)
El Jurásico Medio y Superior constituyó la «edad de oro» de los saurópodos, con una extraordinaria diversificación que dio origen a los principales linajes:
- Aparición de diplodócidos como Diplodocus y Apatosaurus.
- Desarrollo de los braquiosáuridos como Brachiosaurus y Giraffatitan.
- Surgimiento de los camarasáuridos y otros grupos de saurópodos no titanosaurios.
Durante esta fase, los saurópodos alcanzaron una diversidad morfológica sin precedentes y colonizaron virtualmente todos los continentes, aunque con cierto provincialismo faunístico entre Laurasia y Gondwana.
Fase final (Cretácico: 145-66 millones de años)
Contrariamente a lo que se pensaba inicialmente, los saurópodos no experimentaron un declive generalizado durante el Cretácico, sino una reconfiguración ecológica y taxonómica:
- Disminución o extinción de diplodócidos y braquiosáuridos en muchas regiones.
- Extraordinaria radiación de los titanosaurios, especialmente en Gondwana.
- Desarrollo de linajes especializados como los rebaquisáuridos.
Los titanosaurios, en particular, se convirtieron en los saurópodos dominantes del Cretácico, alcanzando su máxima diversidad durante el Cretácico Superior, poco antes de la extinción que marcaría el fin del Mesozoico. El registro fósil indica que los saurópodos permanecieron como componentes significativos de los ecosistemas terrestres hasta el mismo final del período Cretácico.
Distribución paleobiogeográfica
La distribución global de los sauropodomorfos estuvo profundamente influenciada por la configuración cambiante de los continentes a lo largo del Mesozoico y por factores paleoecológicos regionales:
Pangea unificada (Triásico Superior)
Durante sus primeras etapas evolutivas, los sauropodomorfos se beneficiaron de la conexión terrestre que proporcionaba el supercontinente Pangea, permitiéndoles dispersarse ampliamente:
- Sudamérica: Albergó algunos de los primeros sauropodomorfos conocidos, como Saturnalia y Unaysaurus en Brasil, y Riojasaurus y Coloradisaurus en Argentina.
- Europa: Destacan los extraordinarios yacimientos de Plateosaurus en Alemania y Suiza, que han proporcionado cientos de especímenes.
- Sudáfrica: La formación Elliot ha proporcionado numerosos fósiles de sauropodomorfos basales como Massospondylus y Antetonitrus.
- Asia: Formas como Lufengosaurus y Yunnanosaurus en China demuestran la amplia distribución del grupo.
- Norteamérica: Aunque con registro más limitado, taxones como Anchisaurus evidencian su presencia.
Fragmentación inicial (Jurásico Inferior y Medio)
A medida que Pangea comenzaba a fragmentarse, empezaron a surgir patrones de endemismo regional:
- El corredor hispano-marroquí en el Tetis occidental facilitó intercambios faunísticos entre Europa y África del Norte.
- Las conexiones intermitentes entre Asia y Norteamérica a través del puente de Beringia permitieron dispersiones bidireccionales.
- El progresivo aislamiento de Gondwana comenzó a generar linajes distintivos en los continentes del sur.
Provincialismo acentuado (Jurásico Superior y Cretácico)
La creciente separación continental generó patrones biogeográficos más complejos:
- Laurasia (Norteamérica y Eurasia): Dominada por diplodócidos, camarasáuridos y braquiosáuridos durante el Jurásico Superior, con disminución de la diversidad de saurópodos en Norteamérica durante el Cretácico (a excepción del «Alamosaurus event» del Maastrichtiense).
- Gondwana occidental (Sudamérica y África): Desarrollo de linajes endémicos como los rebaquisáuridos y posteriormente una extraordinaria radiación de titanosaurios, especialmente en Sudamérica.
- Gondwana oriental (Madagascar, India, Australia, Antártida): Registro más fragmentario, pero con presencia notable de titanosaurios y otros grupos gondwánicos.
Casos regionales notables
Algunos ejemplos regionales merecen especial atención:
- Patagonia argentina: Representa probablemente el hotspot evolutivo más importante para los titanosaurios, con una extraordinaria diversidad que incluye algunos de los dinosaurios más grandes conocidos como Argentinosaurus, Patagotitan y Puertasaurus.
- Formación Morrison (EE.UU.): Este ecosistema del Jurásico Superior albergó una notable diversidad de saurópodos coexistentes, incluyendo Diplodocus, Apatosaurus, Barosaurus, Camarasaurus y Brachiosaurus, demostrando una sofisticada partición de recursos ecológicos.
- Asia central y China: La relativa continuidad de su registro fósil ha permitido documentar linajes únicos como los mamenchisáuridos, caracterizados por sus cuellos extremadamente largos.
Patrones paleoecológicos
La distribución de los sauropodomorfos no solo estuvo determinada por barreras geográficas, sino también por factores ecológicos:
- Preferencias ambientales: Aunque ocuparon diversos hábitats, los estudios tafonómicos y sedimentológicos sugieren que muchos saurópodos preferían ambientes riparios (cercanos a cursos de agua) y llanuras de inundación.
- Gradientes latitudinales: Su distribución abarcaba desde regiones ecuatoriales hasta latitudes polares, como evidencian los hallazgos en la Antártida, demostrando una notable tolerancia climática.
- Respuestas a cambios climáticos: Existe evidencia de migraciones estacionales en algunas poblaciones, posiblemente relacionadas con la disponibilidad de recursos alimentarios.
- Patrones de extinción regional: Algunas regiones experimentaron extinciones locales de determinados linajes mucho antes de la extinción global del final del Cretácico, sugiriendo vulnerabilidades específicas ante cambios ecológicos regionales.
La extraordinaria distribución global y longevidad temporal de los sauropodomorfos demuestra su notable capacidad adaptativa y resiliencia evolutiva. A lo largo de más de 150 millones de años, estos dinosaurios no solo sobrevivieron a múltiples eventos de extinción, sino que continuaron diversificándose y colonizando nuevos territorios, consolidándose como uno de los grupos de vertebrados terrestres más exitosos de la historia de nuestro planeta.

Importancia paleontológica y descubrimientos relevantes
La investigación sobre los sauropodomorfos ha contribuido significativamente a nuestra comprensión de la evolución, paleobiología y paleoecología de los dinosaurios y, en un sentido más amplio, a nuestro conocimiento sobre la historia de la vida en la Tierra. A continuación, exploraremos la importancia paleontológica de este grupo y algunos de los descubrimientos más relevantes que han transformado nuestra visión sobre estos fascinantes animales.
Contribuciones al conocimiento paleontológico
Evolución del gigantismo
Los saurópodos representan el caso más extremo de gigantismo terrestre conocido, lo que los convierte en modelos ideales para estudiar:
- Los límites biomecánicos del tamaño corporal en animales terrestres.
- Las adaptaciones fisiológicas necesarias para sostener organismos de gran tamaño.
- Los factores evolutivos y ecológicos que favorecen el gigantismo.
Investigadores como Martin Sander y su equipo han desarrollado el concepto de «factores de posibilitación» (enabling factors) para explicar cómo múltiples innovaciones anatómicas, fisiológicas y ecológicas se combinaron para permitir el gigantismo extremo en saurópodos, ofreciendo un fascinante modelo de macroevolución.
Sistemas respiratorios y metabolismo
El estudio de la neumatización del esqueleto en saurópodos ha proporcionado valiosa información sobre:
- La evolución del sistema respiratorio tipo aviano, con importantes implicaciones para entender la fisiología de los dinosaurios no avianos.
- Los patrones metabólicos de animales extintos.
- La relación entre tamaño corporal y demandas energéticas.
Estos estudios han contribuido a reconsiderar la visión tradicional de los dinosaurios como «reptiles gigantes», reconociéndolos como organismos con fisiologías complejas y posiblemente con tasas metabólicas elevadas.
Paleoecología de ecosistemas mesozoicos
Como megaherbívoros dominantes, los sauropodomorfos jugaron un papel ecológico fundamental:
- Modificación del paisaje vegetal a través del consumo selectivo y no selectivo.
- Dispersión de semillas a larga distancia.
- Reciclaje de nutrientes a gran escala.
El estudio de las interacciones entre saurópodos y las comunidades vegetales mesozoicas ha permitido reconstruir complejas redes tróficas y entender mejor la coevolución entre dinosaurios herbívoros y plantas durante este período crítico de la historia de la Tierra.
Descubrimientos e hitos en la investigación
Descubrimientos históricos tempranos
- 1825: Publicación de Megalosaurus por William Buckland, que incluyó material ahora reconocido como perteneciente a un saurópodo.
- 1841: Richard Owen establece el término «Dinosauria», basándose parcialmente en material de sauropodomorfos.
- 1878: Descubrimiento de los primeros esqueletos relativamente completos de saurópodos en Como Bluff (Wyoming), incluyendo especímenes de Apatosaurus y Diplodocus, por Edward Drinker Cope y Othniel Charles Marsh durante la famosa «Guerra de los Huesos».
- 1905: Montaje del primer esqueleto de Diplodocus carnegii (conocido como «Dippy») en el Museo Carnegie de Pittsburgh, del cual se realizarían réplicas enviadas a museos de todo el mundo, popularizando la imagen de los saurópodos globalmente.
Avances significativos del siglo XX
- Expediciones de Janensch a Tendaguru (1909-1913): Resultaron en el descubrimiento de ejemplares extraordinarios de Giraffatitan (anteriormente clasificado como Brachiosaurus) y otros saurópodos en lo que hoy es Tanzania.
- Expediciones del Museo Americano de Historia Natural a la Patagonia (1921-1922): Lideradas por Walter Granger, documentaron la diversidad de saurópodos sudamericanos.
- Descubrimiento de nidos y huevos en Auca Mahuevo, Argentina (1997): Proporcionaron la primera evidencia inequívoca sobre la reproducción de saurópodos, incluyendo embriones preservados.
- Hallazgo de Argentinosaurus (1987) y su descripción formal en 1993 por José Bonaparte y Rodolfo Coria, estableciendo nuevos récords de tamaño para dinosaurios terrestres.
Descubrimientos tecientes transformadores
- Titanosaurios gigantes de la Patagonia: El descubrimiento y descripción de Patagotitan mayorum en 2017, basado en múltiples individuos excepcionalmente completos, ha proporcionado datos cruciales sobre el límite superior del tamaño corporal en dinosaurios.
- Sauropodomorfos basales completos: Hallazgos como el excepcional material de Mussaurus patagonicus, incluyendo individuos de todas las etapas ontogenéticas, han revolucionado nuestra comprensión sobre el desarrollo y evolución temprana del grupo.
- Aplicación de nuevas tecnologías: El uso de tomografía computarizada, análisis biomecánicos computerizados, histología ósea y geoquímica isotópica ha permitido investigaciones sin precedentes sobre la biología de los sauropodomorfos.
- Evidencia de tejidos blandos: Descubrimientos excepcionales como impresiones de piel de titanosaurios en Argentina o la preservación de estructuras queratinosas asociadas a espinas dérmicas.
Yacimientos paleontológicos excepcionales
Algunos yacimientos han sido particularmente importantes para avanzar en nuestro conocimiento sobre los sauropodomorfos:
Europa
- Trossingen y Halberstadt (Alemania): Han proporcionado cientos de especímenes de Plateosaurus, permitiendo estudios detallados sobre variación individual y ontogenia.
- Lo Hueco (España): Yacimiento cretácico que ha revelado una extraordinaria concentración de titanosaurios, incluyendo especímenes articulados y asociados.
América del Norte
- Formación Morrison: Este extraordinario conjunto de yacimientos del Jurásico Superior ha proporcionado la mayoría de los esqueletos montados de saurópodos en museos de todo el mundo, incluyendo géneros icónicos como Diplodocus, Apatosaurus y Brachiosaurus.
- Cantera Howe (Big Bend, Texas): Descubierta en la década de 1930, alberga una extraordinaria concentración de fósiles del titanosaurio Alamosaurus, proporcionando información crucial sobre la recolonización de Norteamérica por saurópodos durante el Cretácico tardío.
Sudamérica
- Formación Neuquén (Argentina): Una de las regiones más productivas para el estudio de titanosaurios, incluyendo los hallazgos de Argentinosaurus, Puertasaurus y más recientemente Patagotitan.
- Auca Mahuevo (Argentina): El mayor sitio de nidificación de dinosaurios conocido, ha proporcionado información sin precedentes sobre la reproducción de los titanosaurios, incluyendo huevos con embriones preservados.
- Formación Laguna Colorada (Argentina): Ha proporcionado el extraordinario material de Mussaurus, incluyendo grupos familiares completos que sugieren comportamiento social complejo en sauropodomorfos basales.
África
- Tendaguru (Tanzania): Comparable en importancia a la Formación Morrison, este yacimiento del Jurásico Superior ha proporcionado numerosos especímenes de saurópodos, incluyendo el famoso Giraffatitan.
- Formación Elliot (Sudáfrica y Lesotho): Fundamental para comprender la evolución temprana de los sauropodomorfos, con numerosos especímenes de taxones como Massospondylus, Antetonitrus y Lessemsaurus.
- Kem Kem (Marruecos): Ha revelado la presencia de rebaquisáuridos especializados como Rebbachisaurus y el extraordinario Nigersaurus.
Asia
- Formación Dashanpu (China): Ha proporcionado esqueletos excepcionalmente completos de mamenchisáuridos, saurópodos con cuellos extremadamente largos característicos de Asia durante el Jurásico.
- Desierto de Gobi (Mongolia y China): Aunque más conocido por sus terópodos y ceratopsianos, también ha revelado importante material de titanosaurios asiáticos.

Impacto cultural y educativo
Los sauropodomorfos, particularmente los saurópodos, han tenido un impacto cultural enorme:
- Se han convertido en iconos culturales reconocibles globalmente, apareciendo en innumerables representaciones artísticas, películas, documentales y literatura.
- Han estimulado la imaginación pública como pocos otros animales extintos, ayudando a popularizar la paleontología.
- Esqueletos montados como el famoso Diplodocus («Dippy») del Museo de Historia Natural de Londres o el Giraffatitan del Museo de Historia Natural de Berlín se han convertido en símbolos institucionales y atracciones turísticas.
- Han proporcionado importantes herramientas educativas para explicar conceptos evolutivos, anatómicos y ecológicos complejos.
El estudio de los sauropodomorfos continúa siendo un campo extraordinariamente dinámico de la paleontología, donde nuevos descubrimientos y metodologías analíticas permiten responder preguntas fundamentales sobre la evolución y la paleobiología, al tiempo que generan nuevas incógnitas. Su importancia trasciende la paleontología, contribuyendo a nuestra comprensión más amplia de los procesos evolutivos y los límites de la biología animal.
Conclusiones: El legado de los gigantes
Los sauropodomorfos representan uno de los capítulos más fascinantes en la historia evolutiva de los vertebrados terrestres. Su extraordinario recorrido, desde modestos bípedos del Triásico hasta los colosos cuadrúpedos del Jurásico y Cretácico, nos ofrece una ventana privilegiada para comprender los procesos macroevolutivos y las complejas interacciones entre forma, función y ecología a lo largo del tiempo geológico.
Lecciones evolutivas
El estudio de los sauropodomorfos nos ha proporcionado valiosas lecciones sobre la evolución:
- Experimentación morfológica: La historia evolutiva del grupo demuestra cómo las innovaciones anatómicas pueden abrir nuevos nichos ecológicos y posibilitar trayectorias evolutivas inesperadas.
- Contingencia histórica: Su evolución ilustra cómo eventos históricos contingentes, como extinciones y oportunidades ecológicas, pueden influir profundamente en las trayectorias evolutivas.
- Restricciones y oportunidades: El gigantismo de los saurópodos representa un fascinante caso de estudio sobre cómo las leyes físicas imponen limitaciones, pero también cómo la selección natural puede encontrar soluciones innovadoras dentro de esas restricciones.
- Éxito evolutivo: Su persistencia durante más de 150 millones de años demuestra que el éxito evolutivo no siempre requiere complejidad cognitiva o versatilidad ecológica, sino adaptaciones eficientes a nichos específicos.
Relevancia contemporánea
Más allá de su importancia histórica, el estudio de los sauropodomorfos tiene relevancia para cuestiones científicas actuales:
- Límites biofísicos: La investigación sobre el gigantismo en saurópodos contribuye a nuestra comprensión de los límites físicos y fisiológicos del tamaño corporal en animales terrestres, con posibles aplicaciones en biología comparada y biomedicina.
- Cambio climático: Como megaherbívoros que sobrevivieron a múltiples cambios climáticos globales, ofrecen perspectivas sobre la resiliencia y vulnerabilidad de los grandes vertebrados ante fluctuaciones ambientales.
- Interacciones planta-herbívoro: Su coevolución con las comunidades vegetales mesozoicas proporciona modelos para entender las complejas dinámicas entre plantas y grandes herbívoros en ecosistemas actuales.
- Métodos interdisciplinarios: El estudio de sauropodomorfos ha estimulado avances metodológicos en paleobiología, biomecánica, paleohistología y reconstrucción paleoecológica que benefician a múltiples campos científicos.
Preguntas pendientes y horizontes futuros
A pesar de los significativos avances en nuestra comprensión de los sauropodomorfos, numerosas preguntas continúan estimulando la investigación:
- ¿Cuáles fueron los límites precisos del tamaño corporal que podían alcanzar y qué factores los determinaban?
- ¿Cómo eran exactamente sus sistemas digestivos y metabolismo, y cómo evolucionaron estas características a lo largo del tiempo?
- ¿Qué grado de complejidad social y comportamental exhibían realmente?
- ¿Cómo influyeron las condiciones paleoambientales cambiantes en su evolución y distribución?
- ¿Qué papel jugaron en la estructuración de los ecosistemas mesozoicos y cómo respondieron a los cambios en las comunidades vegetales?
El horizonte investigador promete emocionantes avances a través de:
- Nuevos descubrimientos fósiles en regiones poco exploradas
- Aplicación de tecnologías emergentes como análisis genómicos de ADN antiguo en parientes avianos
- Modelos computacionales más sofisticados para simulaciones biomecánicas y ecológicas
- Enfoques interdisciplinarios que integren datos paleontológicos, geológicos y biológicos
Reflexión final
Los sauropodomorfos no son simplemente curiosidades prehistóricas, sino extraordinarios ejemplos de las posibilidades y límites de la evolución biológica. Su estudio nos recuerda la profunda historicidad de la vida en nuestro planeta y cómo formas tan diferentes a cualquier animal actual pudieron dominar los ecosistemas terrestres durante un período mucho más largo que el que los humanos han existido.
El legado de estos gigantes perdura no solo en los fósiles que dejaron atrás, sino en la inspiración científica que continúan proporcionando y en cómo han transformado nuestra comprensión del mundo natural y sus posibilidades. Como escribió el paleontólogo Stephen Jay Gould, «la vida en la Tierra ha evolucionado como una narrativa de contingencia histórica», y pocos capítulos de esa narrativa son tan fascinantes como el de los sauropodomorfos.
Al contemplar un esqueleto reconstruido de un saurópodo en un museo, o al imaginar estos colosos recorriendo los paisajes mesozoicos, nos conectamos con un pasado remoto pero real, recordándonos que la historia de la vida es mucho más rica, extraña y maravillosa de lo que podríamos haber imaginado.

Referencias bibliográficas
Apesteguía, S. (2005). Evolution of the titanosaur metacarpus. In K. Carpenter & V. Tidwell (Eds.), Thunder-Lizards: The Sauropodomorph Dinosaurs (pp. 321-345). Indiana University Press.
Bates, K. T., Mannion, P. D., Falkingham, P. L., Brusatte, S. L., Hutchinson, J. R., Otero, A., Sellers, W. I., Sullivan, C., Stevens, K. A., & Allen, V. (2016). Temporal and phylogenetic evolution of the sauropod dinosaur body plan. Royal Society Open Science, 3(3), 150636.
Bonaparte, J. F., & Coria, R. A. (1993). Un nuevo y gigantesco saurópodo titanosaurio de la Formación Río Limay (Albiano-Cenomaniano) de la Provincia del Neuquén, Argentina. Ameghiniana, 30(3), 271-282.
Carballido, J. L., Pol, D., Otero, A., Cerda, I. A., Salgado, L., Garrido, A. C., Ramezani, J., Cúneo, N. R., & Krause, J. M. (2017). A new giant titanosaur sheds light on body mass evolution among sauropod dinosaurs. Proceedings of the Royal Society B: Biological Sciences, 284(1860), 20171219.
Cerda, I. A., Salgado, L., & Powell, J. E. (2012). Extreme postcranial pneumaticity in sauropod dinosaurs from South America. Paläontologische Zeitschrift, 86(4), 441-449.
Chiappe, L. M., Salgado, L., & Coria, R. A. (2001). Embryonic skulls of titanosaur sauropod dinosaurs. Science, 293(5539), 2444-2446.
Curry Rogers, K., & Wilson, J. A. (Eds.). (2005). The Sauropods: Evolution and Paleobiology. University of California Press.
Klein, N., Remes, K., Gee, C. T., & Sander, P. M. (Eds.). (2011). Biology of the Sauropod Dinosaurs: Understanding the Life of Giants. Indiana University Press.
Mannion, P. D., Upchurch, P., Barnes, R. N., & Mateus, O. (2013). Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriforms. Zoological Journal of the Linnean Society, 168(1), 98-206.
Otero, A., & Salgado, L. (2015). El registro de Sauropodomorpha (Dinosauria) de la Argentina. Publicación Electrónica de la Asociación Paleontológica Argentina, 15(1), 69-89.
Pol, D., Garrido, A., & Cerda, I. A. (2011). A new sauropodomorph dinosaur from the Early Jurassic of Patagonia and the origin and evolution of the sauropod-type sacrum. PLoS ONE, 6(1), e14572.
Salgado, L., Coria, R. A., & Calvo, J. O. (1997). Evolution of titanosaurid sauropods. I: Phylogenetic analysis based on the postcranial evidence. Ameghiniana, 34(1), 3-32.
Sander, P. M. (2013). An evolutionary cascade model for sauropod dinosaur gigantism – overview, update and tests. PLoS ONE, 8(10), e78573.
Sander, P. M., Christian, A., Clauss, M., Fechner, R., Gee, C. T., Griebeler, E. M., Gunga, H. C., Hummel, J., Mallison, H., Perry, S. F., Preuschoft, H., Rauhut, O. W. M., Remes, K., Tütken, T., Wings, O., & Witzel, U. (2011). Biology of the sauropod dinosaurs: the evolution of gigantism. Biological Reviews, 86(1), 117-155.
Sereno, P. C. (1999). The evolution of dinosaurs. Science, 284(5423), 2137-2147.
Upchurch, P., Barrett, P. M., & Dodson, P. (2004). Sauropoda. In D. B. Weishampel, P. Dodson, & H. Osmólska (Eds.), The Dinosauria (2nd ed., pp. 259-322). University of California Press.
Wedel, M. J. (2009). Evidence for bird-like air sacs in saurischian dinosaurs. Journal of Experimental Zoology Part A: Ecological Genetics and Physiology, 311A(8), 611-628.
Wilson, J. A. (2002). Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society, 136(2), 217-276.
Xing, L., McKellar, R. C., Xu, X., Li, G., Bai, M., Persons, W. S., Miyashita, T., Benton, M. J., Zhang, J., Wolfe, A. P., Yi, Q., Tseng, K., Ran, H., & Currie, P. J. (2016). A feathered dinosaur tail with primitive plumage trapped in mid-Cretaceous amber. Current Biology, 26(24), 3352-3360.
Yates, A. M., & Kitching, J. W. (2003). The earliest known sauropod dinosaur and the first steps towards sauropod locomotion. Proceedings of the Royal Society of London. Series B: Biological Sciences, 270(1525), 1753-1758.
